Patentschrift: "Methode zur Generierung selbstorganisierender Prozesse für autonome Mechanismen und Organismen"
US 6172941 (angemeldet: 16 12 1999,
erteilt 2001) siehe: Google Patents !
EP 1146406A1 (angemeldet: 03 12 1999) (letzter Absatz, Seite 36, 37)
Autor: Erich Bieramperl, 4040 Linz, Österreich
Verstreichzeit-Quantifizierung
Autoadaptions-Theorem
Algorithmus des Lebens
Der
neuronale Code
Die Bedeutung des Tetragrammatons "JHWH"
Ein
Schritt zu einer neuen Universal-Theorie?
Abstract
Die Erfindung beschreibt eine Methode zur Generierung von Auto-Adaption und Selbstorganisation in autonomen Einheiten, z.B. Mechanismen und Organismen, bei denen eine Vielzahl von Sensoren angeordnet ist, die in verschiedene Wahrnehmungs-Bereichszonen unterteilt sind. Die zeitlich/räumlichen Zustandsveränderungen zwischen den Einheiten und ihrer physikalischen Umgebung produzieren Signalamplituden, wobei die aktuellen Verstreichzeiten zwischen den Phasendurchgängen proportional zur jeweiligen Relativgeschwindigkeit quantisiert, registriert, und fortgesetzt mit früheren Verstreichzeiten verglichen werden. Aus entdeckten kovarianten Zeitsequenzen und Zeitmustern erkennt der Mechanismus oder Organismus seinen eigenen Bewegungsverlauf und kann mittels kovarianzgeregelten Energieimpulsen dazu verhalten werden, stets jenen Zustandsveränderungen nachzueifern, für welche die entdeckten kovarianten Zeitsequenzen und Zeitmuster signifikant sind.
Zusammenfassung
Die Erfindung beschreibt eine Methode zur Generierung
selbstorganisierender Prozesse für Prozesse für autonome Systeme. Sie
erstreckt sich sowohl auf den artifiziellen mechanistischen Bereich als
auch auf Systeme molekular/ biologischer und anderer Art. Mittels der
beschriebenen Erfindung ist es möglich, externe Ereignisse mit variablem
Bewegungsverlauf aus subjektiver Sicht des Systems in Echtzeit zu
identifizieren, Zustandsveränderungen zu erkennen, Eigenbewegungen
nachzuvollziehen und zu optimieren, sowie redundanzarme Prozesse zur
Selbstorganisation herzuleiten. Robotik-Systeme der üblichen
stationären Art stützen sich hauptsächlich auf deterministische
weg-abhängige Regelungs- oder Steuerungsprozesse. Die Regelgrößen oder
Stellwerte zu jeder Position sind zumeist im Programmspeicher eines
Computers enthalten. Durch Anordnung mehrerer Wegbezugsebenen können
verschiedene Freiheitsgrade festgelegt werden. Als Wegaufnehmer dienen
Tacho- Geber, Encoder oder Strichcode-Lineale, die von optischen Sensoren
abgetastet werden und Wegstrecken-Zählimpulse liefern. Der Antrieb
erfolgt zumeist mittels Schrittmotore. Es ist auch bekannt, in
wegabhängigen Programmsteuerungen zusätzliche adaptive Regelungsprozesse
vorzusehen, welche auf Daten basieren, die nach der SHANNON-
Quantisierungsmethode (auch "Sampling" genannt) aus den Amplituden von
Sensor- bzw. Messfühler- Signalen gewonnen werden. Sie dienen zur
Korrektur und Modifikation des Ist-Zustandes. Ein fortgesetzter Vergleich
von Soll- und Istwerten ist erforderlich zur Korrektur und Modifikation
des Regelungs-Prozesses. Die errechneten neuen Regel-Parameter werden
dann in den Programmspeicher übernommen. Diese Art von adaptiven
Regelungen ist beispielsweise notwendig, um bei einem Handhabungs-Roboter
Abweichungen vom festgelegten Bewegungsverlauf auszugleichen, der durch
unterschiedliche Lastzustände entsteht. Wenn ein Roboter oder ein
roboter- gesteuertes Vehikel in dieser Weise in einen autonomen Status
versetzt werden soll, so ist es aber in den meisten Fällen unmöglich, die
genaue Position (oder Koordinaten) durch Tacho- oder Encoderimpulse zu
bestimmen. Regelgrößen oder Steuerbefehle können daher nicht exakt vom
Computer ausgegeben bzw. im Rechner vorprogrammiert werden. Dies betrifft
nicht nur roboter- gesteuerte Autos, gleitende Vehikel, Hovercrafts oder
Flugzeuge aller Art, sondern auch schienengebundene Fahrzeuge, wo die
Ableitung von Wegstrecken- Zählimpulsen oftmals ungenau und nicht
reproduzierfähig ist. Verursacht wird dies durch Unebenheiten, Radschlupf
oder Radabnützung. Erkundungs- oder Handhabungsroboter, die etwa zur
Ortung oder Bergung von Gegenständen an schwer zugänglichen oder
gefährlichen Stellen eingesetzt werden, müssen daher manuell oder mit
Computer-Unterstützung ferngesteuert werden. In solchen Fällen sind
Daten- oder Bildübertragungssysteme erforderlich, um auf einem Monitor
die Bewegungsvorgänge des Roboters überwachen zu können. Bei vielen
Roboter- Anwendungen ist dies jedoch nicht sinnvoll. Ein
roboter-gesteuertes Automobil sollte beispielsweise imstande sein, ohne
menschliche Intervention Objekte in Echtzeit zu erkennen,
Gefahrensituationen auszuweichen und seine Geschwindigkeit an die
Umweltsituation anzupassen. In solchen Fällen ist es notwendig, dass der
Bordcomputer die Situation vor Ort erkennen und die auszuführenden
Schritte errechnen kann. Somit sollte das roboter-gesteuerte Vehikel über
ein gewisses Maß an Fähigkeit zur Selbstorganisation verfügen. Dieselben
Erfordernisse gelten auch für andere Robotik-Systeme.
Zwar ist es in
bezug auf autonome mobile Robotersysteme auch bekannt, das ambiente
Umfeld mittels Sensoren abzutasten und die durch zeitdiskrete
Quantisierung (s. Fig. 1) erhaltenen digitalen Sampling- Daten im
Bordcomputer zu analysieren; und es existieren auch statistische
Rechenverfahren und Algorithmen, um die entsprechenden Regelparameter
bereitzustellen. Statistische Methoden zur Abwicklung solcher
Regelsysteme wurden bereits 1949 von Norbert WIENER beschrieben. Nach dem
bekannten SHANNON-Theorem hat hierbei die Abtastung des Umfeldes mit
mindestens der zweifachen Frequenz der Signalamplituden-Bandbreite zu
erfolgen. Damit bleibt ihr Informationsbetrag erhalten. Um die
Eigenbewegung erfassen zu können, sind sehr hohe Abtastraten erforderlich.
Diese weit verbreitete Amplituden-Quantisierungsmethode erfordert die
Korrelation erfasster Messdaten zu jeweiligen vom Program Counter fix
vorgegebenen Zeitpunkten (Ts). Demnach kann dies als deterministischer
Prozess verstanden werden. Die Praxis hat jedoch gezeigt, dass selbst bei
Verwendung sehr schneller Prozessoren und höchster Samplingraten keine
ausreichende Effizienz erreicht werden kann. Die Zahl redundanter Daten
und Rechenoperationen steigt drastisch an, wenn ein sensorgesteuertes
Vehikel auf neue Hindernisse trifft oder sich mit variabler
Geschwindigkeit auf neue Umwelt- Gegebenheiten adaptieren soll.
Tatsächlich lässt Claude Shannons Quantisierungsmethode keine Erkennung
eines analogen Signalamplituden-Verlaufes in Echtzeit zu; insbes. bei
wechselnden physikalischen Zuständen oder variablen Bewegungsverläufen,
welche die Erfassung zusätzlicher Information über die
Momentan-Geschwindigkeit erforderlich macht. Dies gilt grundsätzlich auch
bei Verwendung von Laser- oder Ultraschallsensoren, bei denen
Entfernungsdaten digital erfasst und verarbeitet werden. Daher eignet sich
diese Quantisierungsmethode zwar dazu, den Spurverlauf einer
Eigenbewegung zu rekonstruieren, wie dies in
Pat.
AT 397 869 beschrieben ist, jedoch kaum zur Erkennung der
Eigenbewegung des Roboters oder zu deren auto-adaptiven Reproduzierung.
Manche autonome mobile Robotersysteme arbeiten mit CCD- Kameras und
OCR-Software (Bildanalyse-Verfahren). Sie schließen aus Farbkontrasten
und Helligkeitsunterschieden auf Konturen oder Objekte, die vom Computer
als anzusteuernde Ziele oder als Orientierungs- Merkmale interpretiert
werden. Beispiele dafür sind rechnergestützte Leit- und Lenksysteme,
welche die automatische Nachführung von Fahrzeugen durch Mittellinien,
Seitenplanken oder Straßenränder ermöglichen. CCD- Sensoren sind - wenn
man ihre Arbeitsweise betrachtet - Analogspeicher, die nach dem bekannten
Eimerkettenprinzip funktionieren. Auf einem MOS- Siliziumchip befinden
sich dichtgepackte Kondensatoren, die durch den fotoelektrischen Effekt
auf ein bestimmtes Potential aufgeladen werden. Jedes Ladungspaket
entspricht einem bestimmten Bildpunkt, genannt "Pixel", wobei die Ladung
jedes Pixels Aufschluss über die Helligkeit an der jeweiligen Stelle des
Bildes gibt. Durch Anlegen von Steuerspannungs- Frequenz werden die
Ladungspakete Pixel für Pixel über das CCD verschoben, wo sie am Output
als serielle Videosignale erscheinen. Um sie im Computer zu verarbeiten,
müssen sie in digitale Größen umgewandelt werden. Ihre Verarbeitung
erfordert eine hohe Anzahl von redundanten Daten und Rechenprozessen,
weshalb die digitale Erfassung von längeren Sequenzen elektronische
Speicher mit extrem großer Kapazität erfordert. Eine Erkennung isomorpher
Sequenzen in Bewegungsverläufen ist nur mit großem Aufwand und in
zeitlicher Versetzung möglich, weshalb derartige auf CCD- Sensoren
basierende Robotiksysteme nur in sehr beschränkter Weise zur
auto-adaptiven Nachvollziehung von Eigenbewegungen geeignet sind. Bei
jeder Wiederholung desselben Bewegungsvorganges an gleicher Strecke
müssen alle Regelparameter mittels Bildanalyse neu berechnet werden.
Verändern sich Umweltbedingungen durch Nebel, Dunkelheit oder Schneefall,
sind solche Systeme zumeist überfordert und nicht einsetzbar.
Pat. AT 400 028
beschreibt ein System zur adaptiven Regelung eines motorgetriebenen
Vehikels, wobei Markierungen oder Signalquellen entlang der Strecke
angeordnet sind, um als Wegweiser zur Einhaltung eines Fahrplanes zu
dienen. Auch Positionen, die durch GPS- Signale ermittelt werden, können
dazu dienen. Beim Passieren dieser Quellen misst das bordseitige
sensor-gekoppelte Computersystem die benötigten Zeiten für jeden
zurückgelegten Streckenabschnitt mittels der in
Pat.
U.S. 4,245.334 beschriebenen Weise der Zeitquantisierung durch erste
und zweite Sensorsignale. Die auf diese Art erworbenen Daten dienen als
Referenzbasis für die Errechnung jener Regelparameter, welche die
Antriebs- und Bremszyklen des Vehikels steuern. Das System arbeitet mit
einer geringen Zahl redundanter Daten, korrigiert sich in auto-adaptiver
Weise selbst, und ist imstande, eine elektronische "Marschtabelle"
nachzuvollziehen. Es eignet sich insbes. zur Einhaltung von
Streckenfahrplänen im Schienenverkehr. Allerdings ist es mit diesem System
nicht möglich, ambiente Objekte Umgebungen festzustellen. Es ist Aufgabe
dieser Erfindung, eine umfassende Methode zur Generierung autonomer
selbst-organisierender Robotik-Systeme bereitzustellen, die es
ermöglicht, externe Signale, Objekte, Ereignisse, physikalische Zustände
oder Umgebungen aus subjektiver Sicht in Echtzeit zu identifizieren.
Dadurch wird es ermöglicht, die Muster von Eigenbewegungen zu erkennen
und die eigene Verhaltensweise in autoadaptiver Weise zu reproduzieren und
zu optimieren. Es ist eine weitere Aufgabe der Erfindung, mittels dieser
Methode einen autonomen Trainingsroboter für den Einsatz im Sport
bereitzustellen, der imstande ist, einen vom Benutzer vorgegebenen
Bewegungsverlauf zu identifizieren, zu reproduzieren und zu optimieren;
außerdem die ideale Spur und Tempoverteilung selbsttätig zu suchen,
bestimmte Abstände und Zeittabellen einzuhalten, Eigenbewegungs-Verläufe,
Geschwindigkeiten, Rundenzeiten, Teilzeiten und Gesamtzeiten auf einem
Monitor darzustellen; sowie die erfassten Daten optisch oder akustisch
auszugeben.
Die gestellten Aufgaben werden bei der erfindungsgemäßen Methode im
Allgemeinen durch Anordnung von Sensoren oder Rezeptoren an dem sich
bewegenden System (z.B. einem autonomen Robotik-System) gelöst, welche
die ambienten Signalquellen, deren Amplituden durch Festlegen einer
Anzahl von Schwellwerten unterteilt werden, abtasten. Dadurch werden
Wahrnehmungsbereichszonen geschaffen. Die Verstreichzeiten aller
Phasendurchgänge in allen Zonen werden mittels analoger oder digitaler
STQ-Quantisierung fortgesetzt erfasst, und die Zeitzähltakte in
Abhängigkeit von der relativen Momentan- Geschwindigkeit, die durch die
Phasenverschiebung der Signalverläufe benachbarter gleichartiger Sensoren
bestimmt ist, automatisch moduliert. Die gezählten Impulse korrelieren
annähernd mit den zurückgelegten Distanzwerten (dnnn). Bei dieser Methode
ist die Abtastung von Signalamplituden kein deterministischer Prozess;
sie erfolgt nicht zu vorgegebenen Zeitpunkten in bestimmten Zeittakten.
Die Erfassung, Verarbeitung und Analyse der Verstreichzeiten erfolgt
weitgehend nach probabilistischen Prinzipien. Daraus resultiert die
physikalisch bedeutsame Tatsache, dass die Parameter zur Beschreibung der
externen Umgebung vom System eigentlich nicht "objektiv gemessen",
sondern als zeitliche Abfolgen quasi "subjektiv empfunden" werden. Das
System selbst fungiert hierbei als "Beobachter" des Prozesses. In der
technischen Fachliteratur werden Verstreichzeiten (im Zusammenhang mit
deterministischem Timing) auch als "Signal- Laufzeiten" oder
"Zeitintervalle" bezeichnet. Bei der beschriebenen Erfindung erfolgt die
Quantisierung der sogenannten "STQ-Verstreichzeiten" im Verlauf des
Signalerkennungs- Prozesses durch Starten und Stoppen einer Vielzahl von
Zeitmessern an allen Phasen- Übergängen. Sie ergeben zusammen einen Strom
von Zeitdaten. Jede Verstreichzeit zwischen Phasendurchgängen in der
gleichen Wahrnehmungsbereichszone kann ebenso erfasst werden wie die
Verstreichzeiten zwischen den Phasendurchgängen von einem niedrigen zu
einem höheren Schwellwert und umgekehrt. Man unterscheidet grundsätzlich
drei verschiedene Arten von STQ-Quantisierung (bzw.
Verstreichzeitmessung):
....ist die Verstreichzeit, festgelegt durch die beim Vorbeibewegen eines
ersten Sensors bzw. Rezeptors S2 und eines gleichgearteten
zweiten Sensors bzw. Rezeptors S entlang einer korrespondierenden
externen Signalquelle Q hervorgerufene Signal- amplitude. Sie
wird von der steigenden Flanke beim Phasendurchgang iTv1.1 des
ersten Sensorsignals zur steigenden Flanke beim Phasendurchgang iTw1.1 des
zweiten Sensorsignals, desgl. von iTv2.1 zu iTw2.1, von iTv3.1 zu
iTw3.1 usw. gemessen., wobei sich die Phasendurchgänge auf die
gleichen Schwellwertpotentiale P1,2,3... beziehen. STQ(v)- Zeiten
können auch von fallenden Flanken gemessen werden. Sie dienen als
Parameter für die momentane Relativgeschwindigkeit (vm) des sich
bewegenden Systems.
STQ(i) = sensitivity/time quantum of
interarrival = Tw1,2,3...
....ist die Verstreichzeit,
festgelegt durch die von einem Sensor (bzw. Rezeptor) S1 im Feld
einer korrespondierenden externen Signalquelle Q und/oder durch die beim
Vorbeibewegen eines Sensors bzw. Rezeptors S entlang mehrerer
gleichartiger externer Signalquellen Q1,2,3...hervorgerufene
Signalamplitude. Die Verstreichzeit wird gemessen von der
steigenden Flanke beim Phasendurchgang iTw1.1 des steigenden
Sensorsignals zur fallenden Flanke beim Phasendurchgang iTw1.2 des
fallenden Sensorsignals, desgl. von iTw2.1 zu iTw2.2, von iTw3.1
zu iTw3.2 usw., sowie von der fallenden Flanke beim
Phasendurchgang iTw1.2 weiter zur nächsten steigenden Flanke beim
Phasendurchgang iTw1.3, desgl. von iTw2.2 zu iTw2.3, von iTw3.2 zu
iTw3.3 usw., wobei sich die Phasendurchgänge auf die gleichen
Schwellwert- potentiale P1,2,3....beziehen. Wird die
Zeitzählfrequenz für die Messung der STQ(i)- Verstreichzeiten Tw(1,2,3...)
proportional zur momentanen Relativgeschwindigkeit vm (welche
mittels STQ(v)- Parameter erfasst ist) erhöht oder verringert, so
haben die gezählten Impulse korrelativen Bezug zur Strecke, die
das sensor-gekoppelte System zurückgelegt. Es ist klar
ersichtlich, dass die solchermaßen adaptierten Verstreichzeiten nicht
ident mit realen physischen Zeiten sind, die von üblichen Zeitmessern
für jene relative Strecke gemessen würden. Besteht
jedoch absolute physikalische Invarianz zwischen dem sich bewegenden
System und seiner Umgebung (z. B. Synchronität), so kann kein
STQ-Parameter erfasst werden.
STQ(d) =
sensitivity/time quantum of differentiation = Td1,2,3...
....ist die Verstreichzeit, festgelegt durch die von einem Sensor (bzw.
Rezeptor) S1 im Bereich einer korrespondierenden externen
Signalquelle Q1,2,3.. hervorgerufene Signalamplitude, gemessen
von der steigenden Flanke beim Phasendurchgang iTw1 des steigenden
Sensor-Signalverlaufes zur steigenden Flanke beim nächsthöheren Phasen-
Durchgang iTw2, von iTw2 zu iTw3, von iTw3 zu iTw4 usw., sowie in
umgekehrter Folge beim fallenden Signalverlauf, wobei sich die
Phasendurchgänge auf die gleichen Schwellwertpotentiale
P1,2,3..beziehen. STQ(d)-Daten stellen Differential- Parameter
für die Flankensteilheit von Signalamplituden dar und somit für deren
Frequenz, außerdem dienen sie der Plausibilitätsprüfung und
Verifizierung von anderen korrespondierenden STQ-
Verstreichzeitdaten. Die Relativbewegung zwischen Sensor und
Signalquelle wird bei dieser Messung nicht berücksichtigt.
Besteht keine Relativbewegung zwischen Sensoren und Quellen (jedoch
Varianz von Feldern) so werden Zustandsveränderungen eines Feldes
aus STQ(i)- und STQ(d)- Verstreichzeiten erfaßt und
identifiziert. Wenn aber absolute physikalische Invarianz gegeben
ist, können keine STQ- Quanten erfasst werden, und Erkennung ist
ausgeschlossen. STQ(v)- Daten werden vorwiegend dazu benötigt, um die
räumliche Umgebung unter relativer Bewegung erkennen und/oder aus
identifizierten Bewegungsprozessen die eigene Bewegung (oder
Teile dieser Bewegung) erkennen sowie in auto-adaptiver Weise
nachvollziehen zu können.
Bei einer im mechanistischen Bereich
angewandten Ausführungsform der erfindungsgemäßen Methode werden die
genannten Wahrnehmungsbereichszonen üblicherweise durch eine Anzahl
elektronischer Schwellwertdetektoren mit unterschiedlich eingestellten
Empfindlichkeitspegel gebildet, und die STQ(i)- und STQ(d)-
Verstreichzeitdaten werden mittels programmierbarer digitaler Timer
erfasst. Der Verstreichzeit- Registrierungsprozess wird an einem iT-
Phasendurchgang sowohl gestartet als auch gestoppt. Dann wird die Zeitdate
in einem Memory gespeichert. Ferner können besagte STQ(v)-
Verstreichzeiten mittels elektronischer Integratoren erfasst werden,
wobei die Aufladezeiten der Kondensatoren jene Spannungs- Potentiale
bestimmen, welche als analoge STQ(v)-Werte an Spannungs/ Frequenz-
Konverter anzulegen sind, um an deren Outputs die digitalen
Zeittaktfrequenzen zur adaptiven Messung der STQ(i)- und STQ(d)-
Verstreichzeiten so zu modulieren, dass sie eine Funktion der
Relativgeschwindigkeit vm bilden.
In einer weiteren,
nicht-mechanistischen Ausführungsform der Erfindung ist vorgesehen, dass
sowohl die genannten Wahrnehmungsbereichszonen bzw. Schwellwertdetektoren
sowie die beschriebenen STQ-Zeitquantisierungsprozesse nicht auf
dieselbe Weise wie analog/ digitale Schaltkreise, sondern auf ähnliche
Art wie molekular/biologisch/chemische Strukturen gebildet sind.
In einer weiteren, allgemeinen Ausführungsform der Erfindung ist
vorgesehen, dass die aktuell erfassten und gespeicherten STQ-
Verstreichzeitdaten fortgesetzt mittels Echtzeit- Analyse mit früher
erfassten und gespeicherten Zeitstrommustern verglichen werden, um
dadurch externe Ereignisse oder physikalische Zustandsveränderungen mit
einem Minimum an Redundanz identifizieren bzw. in Echtzeit erkennen zu
können.
In einer weiteren allgemeinen Ausführungsform der Erfindung
ist vorgesehen, daß mit Sensoren und Einrichtungen zur o.g.
Zeitstrommuster-Erkennung versehene, sich autonom bewegende Systeme, über
Antriebs-, Lenkungs- und Bremsmechanismen verfügen, wobei die Regelung
dieser Mechanismen derart erfolgt, daß das autonom bewegte System (insbes.
ein Robotersystem) zuvor erkannte STQ- Zeitstrommuster in auto-adaptiver
Weise zu reproduzieren vermag. Bei Wiederholung der Bewegung löscht eine
Prozessoreinheit instabile oder ungenügend koordinierte
Tempoverteilungsmuster aus dem Speicher und weist nur jene
Zeitstrommuster als Instruktion zu, welche die Reproduzierung der
Bewegung entlang gleicher Strecke in optimaler koordinierter Weise
gewährleistet. Außerdem ist vorgesehen, die Zeitmessfrequenz für die
genannte STQ(v)-Verstreichzeit- Messung in zeitlichem Maßstab proportional
zu erhöhen oder zu vermindern, wodurch die Tempoverteilung in allen
Bewegungen in selbem Maßstab proportioniert wird. Um die Messungen der
besagten STQ-Verstreichzeitdaten gegebenenfalls zu korrigieren, und um
die Erkennung identer Zeitstrommuster oder die Reproduzierung von
Bewegungs- vorgängen zu verbessern, ist außerdem vorgesehen, die durch
Schwellwertdetektoren gebildeten Wahrnehmungsbereichszonen automatisch
nachzustellen bzw. zu fokussieren, falls es häufige Abweichungen
erfordern. (Dies ist Gegenstand einer weiteren Patent-
anmeldung).
Fig. 1 zeigt ein Diagramm von SHANNON's deterministischer Methode der
zeitdiskreten von Signalamplituden-Verläufen, welche mittels
Analog/Digital-Wandler digitalisiert werden. Im allgemeinen technischen
Sprachgebrauch wird diese Methode auch "Sampling" genannt. Diese
deterministische Methode ist dadurch gekennzeichnet, dass erfasste Daten
(a1, a2...an) mit bestimmten Zeitpunkten (T1, T2, T3,...Tn) korrelieren,
die vom Program Counter eines Prozessors vorgegeben werden.
In der
gegenwärtigen Robotik-Praxis verlangt diese Methode sehr schnelle
Prozessoren, hohe Samplingraten und eine hohe Anzahl redundanter Daten
und Rechenoperationen bei deren Auswertung. Will man auf diese Weise
Daten von Signalamplituden externer Quellen zum Zweck der Information
über die räumliche Umgebung eines Systems, auf welchem ein sensor-
gekoppelter Prozessor installiert ist, erwerben, so lässt SHANNON's
Methode keine ausreichende Gewinnung von Daten über die momentane
Relativgeschwindigkeit und die zeitlich/räumliche Zuordnung zu, die zum
Zweck der Optimierung der Koordinierung der relativen Eigenbewegung
notwendig wären. Eine Erkennung der eigenen Bewegung in Echtzeit ist
daher nicht möglich. Aus diesem Grund ist diese Methode zur Generierung
hocheffizienter autonomer Robotiksysteme zu schwach.
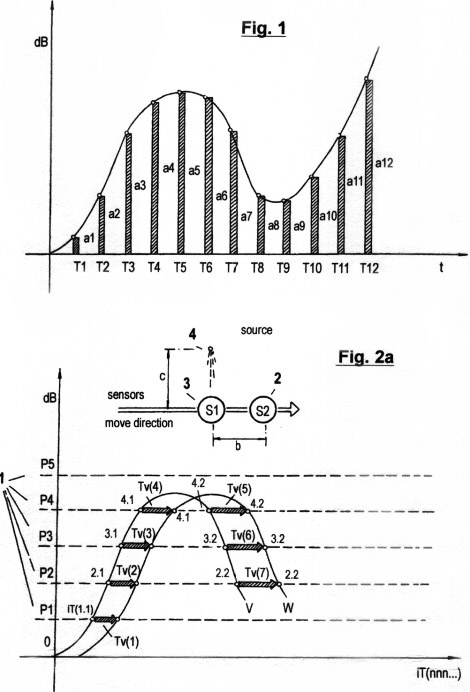
Figs. 2a - c zeigen in drei verschiedene Graphen die sensorische Quantisierung von Signalamplituden verläufen mittels der hierin beschriebenen erfindungsgemäßen Methode. Im Gegensatz zur in Fig. 1 gezeigten Quantisierungsmethode werden hierbei keinerlei "vertikalen Segmente" von Amplitudenverläufen abgetastet; sondern ausschließlich Verstreichzeiten gemessen, und zwar auf drei verschiedene, sich ergänzende Arten. Wie ersichtlich, müssen dazu eine gewisse Anzahl von Schwellwerten 1 (P1, P2...Pn) festgelegt werden, um sensorische Wahrnehmungsbereichzonen zu bilden. Jede Verweildauer und jedes Zeitintervall innerhalb der genannten Zonen wird ebenso erfasst wie die Verstreichzeit zwischen den Durchgängen von einem niederen zu einem höheren Schwellwert und umgekehrt.
Fig. 2a zeigt die erste dieser drei
verschiedenen Arten sensorischer Zeitquantisierung. Sie wird als STQ(v)-
Verstreichzeitmessung (sensitive time quantum - velocity) bezeichnet und
liefert ein Maß für die momentane Relativgeschwindigkeit vm. Man kann sie
auch als Pausendauer zwischen zwei Phasendurchgängen zweier paralleler
Signalverläufe an gleichen Schwellwertpotentialen verstehen. Nicht ganz
korrekt ist dafür die übliche Bezeichnung "Phasenverschiebung". Im Graph
sind die gemessenen STQ(v)-Verstreichzeiten mit Tv(n) bezeichnet. Die
Phasendurchgänge an dem vom Sensor (bzw. vom Receptor) beim Vorbeibewegen
an der entsprechenden externen Signalquelle 4 produzierten
Amplitudenverlauf V sind mit iTv(n.m),
die Phasendurchgänge an dem vom Sensor (bzw. Rezeptor beim Vorbeibewegen
an der gleichen Signalquelle erhaltenen Amplitudenverlauf W mit iTw(n.m)
zeichnet. Unter der idealen Voraussetzung, dass der räumliche Abstand (b)
zwischen den beiden Sensoren 3, 4 im Vergleich zum Abstand c
zwischen Signalquelle und Sensoren minimal ist, dass c beim Vorbeibewegen
konstant bleibt, und dass beide Sensoren (bzw. Rezeptoren) identische
Eigenschaften aufweisen und ein Analogsignal liefern, so werden an den
Outputs der erwähnten Sensoren (bzw. der Sensor-Operationsverstärker) zwei
Signalverläufe V und W produziert,
die annähernd konruent sind. (Abweichungen von der Ideal-Konstellation
können durch autonome Adaption des Sensorsystems in fortschreitend
verbesserter Weise kompensiert werden; dies wird später beschrieben).
Bewegt sich Sensor 2 in der bezeichneten Richtung entlang der
Signalquelle so durchbricht die Signalamplitude V zunächst das definierte
Schwellwertpotential P1 bei Phasendurchgang iTv(1.1). Die ansteigende
Signalflanke startet einen ersten Zeitmesser zur Erfassung der ersten
STQ(v)- Verstreichzeit Tv(1). Die weiter ansteigende Signalamplitude V
durchbricht sodann die Schwellwertpotentiale P2, P3 und P4, wobei bei
jedem Phasendurchgang weitere Zeitmesser zur Erfassung weiterer
Verstreichzeiten Tv(2), Tv(3) und Tv(4) gestartet werden. Inzwischen hat
sich Sensor 3 der Signalquelle 4 genähert und liefert den
Signalamplitudenverlauf W. Durchbricht W bei Phasendurchgang iTw(1.1) das
Schwellwertpotential P1, so stoppt die steigende Flanke den Zeitmesser
zur Erfassung der ersten STQ(v)-Verstreichzeit Tv(1) und speichert sie.
Derselbe Vorgang wiederholt sich für die Verstreichzeiten Tv(2), Tv(3) und
Tv(4) beim Durchbrechen der nächsthöheren Schwellwertpotentiale P2, P3 und P4.
Beginnt die Signalamplitude V zu fallen, so durchbricht sie zunächst den
Schwellwert P4 an der oberen Schulter des Amplitudenverlaufes nach unten.
Nun startet die fallende Flanke einen Zeitmesser zur Erfassung der
nächsten Verstreichzeit Tv(5). Bei den weiteren Phasendurchgängen
iTv(3.2) und iTv(2.2), bei dem die Schwellwerte P3 und P2 nach unten
durchbrochen werden, werden ebenfalls wieder Zeitmesser durch die
fallende Flanke gestartet, um die Verstreichzeiten Tv(6) und Tv(7) zu
messen. Steigt der Verlauf der Signalamplitude V wieder an, so erfolgt
die Erfassung der STQ(v)-Parameter wieder durch die steigende Flanke.
Dasselbe gilt für das Stoppen der besagten Verstreichzeitmesser bei den
Phasen- Durchgängen der parallel zu V verlaufenden Signalamplitude
W.
Fig. 2b zeigt eine weitere Art sensorischer STQ-
Zeitquantisierung. Sie wird als STQ(i)- Verstreichzeitmessung (sensitive
time quantum / interarrival) bezeichnet. Vereinfacht, bedeutet sie die
benötigte Zeit Tw für die Zurücklegung einer relativen Strecke. Man kann
sie auch als Verweildauer eines Signalverlaufes zwischen den
Phasendurchgängen an gleichen Schwellwertpotentialen verstehen. Wird die
Zeitzählfrequenz für ihre Messung proportional zu
Relativgeschwindigkeits-Parameter Tv (bzw. zu STQ(v)- Verstreichzeiten)
beschleunigt oder vermindert, so korrelieren die gezählten und
gespeicherten modulierten Impulse mit der relativen Strecke. Besteht
absolute Invarianz zwischen dem Sensor und den Signalquellen (auch
"Synchronität" genannt), so kann kein STQ(i)-Parameter erfasst werden;
ändert sich jedoch die Signalintensität, so können
STQ(i)-Verstreichzeiten auch dann erhalten werden, wenn keine
Relativbewegung stattfindet. Während einer Bewegung sind sie also nicht
nur zur Erfassung von variablen Signalen erforderlich, sondern auch zum
Abtasten der räumlichen Umgebungen. Im Graph (Fig. 2b) sind die
gemessenen STQ(i)-Verstreichzeiten mit Tw(n) bezeichnet. Die
Phasendurchgänge, welche vom Amplitudenverlauf W produziert werden, wenn
sich Sensor (bzw. Rezeptor) 5 entlang den entsprechenden benachbarten
Signalquel-len 6 und 7 vorbeibewegt, sind mit iTw(n.n) bezeichnet. Sobald
sich Sensor (bzw. Rezeptor) 5 in der bezeichneten Richtung entlang der
Signalquelle 6 bewegt, durchbricht die Signal- Amplitude W zunächst das
Schwellwertpotential P1 bei Phasendurchgang iTw(1.1). Die steigende
Signalflanke startet einen ersten Zeitmesser zur Erfassung der ersten
STQ(i)- Verstreichzeit Tw(1). Danach durchbricht die weiter ansteigende
Signalamplitude W die vorgegebenen Schwellwertpotentiale P2, P3 und P4,
wobei bei jedem Phasendurchgang Zeitmesser gestartet werden, um weitere
Verstreichzeiten Tw(2), Tw(3), Tw(4).... zu erfassen. Inzwischen beginnt
sich Sensor 5 aus dem Bereich der Signalquelle 6 zu entfernen. Die
fallende Signalamplitude W durchbricht zunächst das Schwellwertpotential
P4, worauf beim Phasendurchgang iTw(4.2) nun die fallende Flanke jenen
Zeitmesser stoppt, der zur Erfassung der STQ(i)-Verstreichzeit Tw(4)
gestartet worden war. Gleichzeitig startet dieselbe fallende Flanke einen
weiteren Zeitmesser, der die Verstreichzeit Tw(5) bis zum Eintreffen der
nächsten ansteigenden Flanke misst. Jedoch erscheint diese steigende
Flanke erst beim Vorbeibewegen des Sensors 5 an der entsprechenden
Signalquelle 7. Vorher fällt die Signal- Amplitude W aber bis unter die
Schwellwerte P3 und P2, bei deren Phasendurchgang iTw(3.2) und iTw(2.2)
die Zeitmesser zur Erfassung der STQ(i)- Verstreichzeiten Tw(3.2) und
Tw(2.2) gestoppt werden. Gleichzeitig werden weitere Zeitmesser zur
Erfassung der Verstreichzeiten Tw(6) und Tw(7) gestartet. Sie stoppen
wiederum zu den Phasendurchgängen iT(2.3), iTw(3.3), iTw(4.3) und
iTw(5.1), wenn die Signalamplitude wieder fällt (jedoch nicht vor dem
Vorbeibewegen des Sensors an Signalquelle 7). Nach diesen
Phasendurchgängen starten neue Zeitmesser zur Erfassung der nächsten
Verstreichzeiten Tw(8), Tw(9), Tw(10), Tw(11), usw.
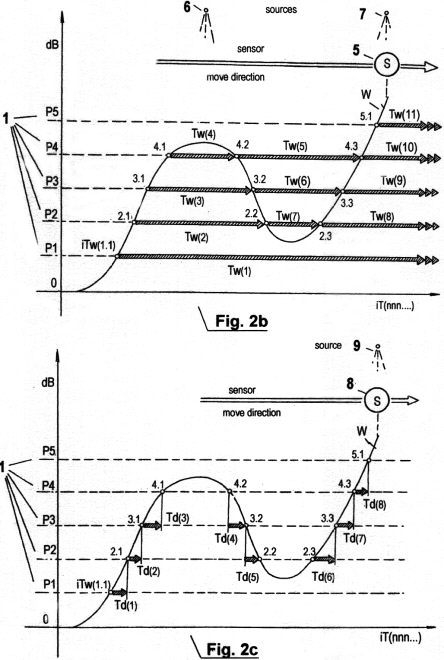
Fig. 2c zeigt
eine dritte Art der sensorischen Zeitquantisierung, die sich von den in
Fig. 2 und Fig. 2b gezeigten Methoden total unterscheidet. Sie wird als
STQ(d)- Verstreichzeit- Messung (sensitive time-quantum - differential)
bezeichnet. Man kann sie auch als Verweildauer Td, gemessen zwischen
einem ersten Phasendurchgang bei einem ersten bestimmten
Schwellwertpotential und einem zweiten Phasendurchgang bei einem zweiten
bestimmten Schwellwertpotential verstehen, das um eine Stufe höher oder
niedriger als das Erstere ist. STQ(d)- Verstreichzeiten sind Parameter
für die Flankensteilheit von Signalamplituden, und somit auch für deren
Frequenz. Mittels schnellem Vergleich von STQ(d)-Verstreichzeiten können
Signalverläufe in Echtzeit erkannt werden. Sie sind daher zur Schaffung
intelligenter Verhaltensweisen ebenso unerläßlich wie STQ(v) oder
STQ(i)-Quanten. Die Quantisierung von STQ(d)- Zeitdaten ist unter allen
physikalischen Zuständen und beliebigen Relativ- Bewegungen zwischen
Sensor und externen Quellen möglich, in welchen STQ(v)- und STQ(i)-
Verstreichzeiten ebenfalls quantisierbar sind. Werden die
STQ(d)-Verstreichzeiten kumulativ und in Serie erfaßt, so können sie zur
Verifizierung und Plausibilitätsprüfung von (gleichfalls erfassten)
STQ(i)-Verstreichzeiten dienen. Im Graph (Fig. 2c) sind die gemessenen
STQ(d)- Verstreichzeiten mit Td(n) bezeichnet. Die Phasendurchgänge,
welche vom Amplitudenverlauf W produziert werden, wenn sich Sensor (bzw.
Rezeptor) 8 im Bereich der Signalquelle 9 befindet, mit iTw(n.n)
bezeichnet. Bewegt sich Sensor 8 in der gezeigten Richtung entlang der
Signalquelle 9, so durchbricht die Signalamplitude W zunächst das
definierte Schwellwert- Potential P1 bei Phasendurchgang iTw(1.1). (Dies
geschieht bei aktivem bzw. variablem Signalquellenfeld selbstverständlich
auch dann, wenn Sensor und korrespondierende Signal- quelle in invarianter
Position zueinander sind). Die steigende Signalflanke startet einen ersten
Zeitmesser zur Erfassung der ersten STQ(d)- Zeitdate. Wenn der weiter
ansteigende Amplitudenverlauf W das nächsthöhere Schwellwertpotential P2
beim Phasendurchgang iTw(2.1) durchbricht, so wird dieser Zeitmesser
gestoppt und die gemessene STQ(d)- Verstreichzeit Td(1) gespeichert.
Gleichzeitig startet der nächste Zeitmesser zur Erfassung und Speicherung
der Verstreichzeit bis zum nächsten Phasendurchgang bei iTw(3.1), wo er
gestoppt wird; dann wird der nächste Timer aktiviert bis zu iTw(4.1), wo
er ebenfalls gestoppt wird usw. Beim Phasendurchgang iTw(4.1) durch
Schwellwertpotential P4 wird der nächste Zeitmesser gestartet. Da aber das
Signal kein nächsthöheres Schwellwertpotential mehr erreicht bevor es
wieder gegen P4 absinkt, kann mit letzterem Zeitmesser keine STQ(d)-
Verstreichzeit erfasst werden. Nur die Quantisierung von
STQ(i)-Verstreichzeiten, wie in Fig. 2b beschrieben, kann in dieser
Situation stattfinden. Die nächste STQ(d)- Verstreichzeit Td(4) wird erst
dann erfasst, wenn die Signalamplitude bei iTw(4.2) unter den Schwellwert
P4 geht, wonach ein weiterer Timer startet, der beim Phasendurchgang
durch den um eine Stufe niedrigeren Schwellwert P3 gestoppt wird.
Gleichzeitig wird der nächste Zeitmesser gestartet usw. In
mechanistischen Anwendungen, wo die Analyse von Signalamplituden die
Quantisierung von STQ(d)-Verstreichzeiten erfordert, werden STQ(d)- Daten
oftmals in Kombination mit STQ(i)-Daten aquiriert. Besteht die Intention
der Anwendung dieser Quantisierungsmethode darin, einem Roboter die
Erkennung seiner Eigenbewegung aus subjektiver Sicht zu ermöglichen
(durch Aufspüren und Abtasten der räumlichen Umgebung beim Bewegen entlang
externer Signalquellen), so werden überwiegend STQ(v)- und STQ(i)-Daten
erfasst. Geht jedoch der Schwerpunkt in Richtung Erkennung instabiler
optischer oder akustischer Quellen, wie etwa Bilder, Musik oder Gespräche
usw., so nimmt der Anteil von STQ(d)-Verstreichzeiten zu, während der
Anteil von STQ(v)-Parameter abnimmt. Findet keinerlei relative Bewegung
statt, so können auch keinerlei Geschwindigkeitsparameter von
irgendwelchen Sensoren abgeleitet werden; nur die Quantisierung von
STQ(d)- und STQ(i)-Verstreichzeiten ist möglich.
Figs. 3 a -
c zeigen einen wichtigen Aspekt der Gestaltung der beschriebenen
Methode im Zusammenhang mit serieller Übertragung von erfaßten
STQ(d)-Verstreichzeiten sowie in Verbindung mit
Zeitzählfrequenzmodulation in Relation zu simultan erfassten STQ(v)-
Parametern, die für die momentane Relativgeschwindigkeit (vm) stehen.
Diese Gestaltungsform der Methode eignet sich allerdings nur dort, wo
überwiegend STQ(d)-Verstreichzeiten gemessen werden; zusammen mit solchen
STQ(i)-Verstreichzeiten (siehe auch Beschreibung zu Fig. 2c), welche bei
den Phasendurchgängen des erreichten maximalen Schwellwerts nahe des
Maximums der Amplitude, oder des erreichten minimalen Schwellwerts nahe
des Minimums der Amplitude, produziert werden. In diesem Fall können alle
gemessenen Verstreichzeiten als serielle Sequenzen dargeboten werden.
Werden hingegen zu jedem Schwellwertpotential STQ(d)- und bzw. nur
STQ(i)-Verstreichzeiten erfasst (siehe Beschreibung zu Fig. 5), so fallen
diese Daten parallel an und müssen auch parallel verarbeitet
werden.
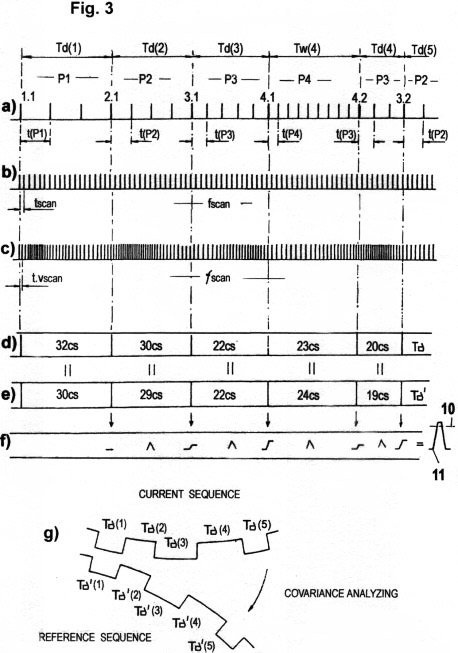
Fig. 3a zeigt, wie eine einfache serielle Impulsfolge zum schnellen Datentransfer der erfassten STQ(d)- Verstreichzeiten ausreicht, wenn die Schwellwertpotentiale P1, P2, P3..., welche jene Phasendurchgänge 1.1, 2.1, 3.1...bestimmen, von denen die STQ-Verstreich- zeiten abgeleitet werden, entweder durch Codes oder durch bestimmte charakteristischen Frequenzen "markiert" sind. Im gezeigten Fall sind diese "Marker" Impulsfolgen mit Perioden t(P1), t(P2), t(P3) ...und Frequenzen f(P1), f(P2), f(P3).... Ihre Modulation erfolgt in Abhängigkeit von den entsprechenden Schwellwertpotentialen. Diese Identifikations-Impulsfolgen (IP) dienen der Identifizierung der bestimmten Schwellwerte P1, P2, P3...(bzw. der Wahrnehmungsbereichszonen 1, 2, 3...). Erst die zusammen mit diesen Identifikations- Impulsen (IP) operierenden invarianten Zeitzählimpulsen (ITPC) mit der Periode tscan (Fig.3b) oder variablen Zeitzählimpulsen (VTPC) mit der Periode t.vscan (s. Fig. 3b, 3c) bewirken die eigentliche Erfassung von STQ(d)-Verstreichzeiten Td(1), Td(2), Td(3), Td(4)... (oder bzw. der STQ(i)- Verstreichzeiten Tw(1), Tw(2), Tw(3), Tw(4)..., welche nahe der Amplituden-Maxima und -Minima auftreten) nach dem bereits beschriebenen Beispiel. Variable VTCP- Impulsfolgen mit der Periode t.vscan, welche automatisch in Relation zu den erfassten STQ(v)-Verstreichzeiten (d.h. zur augenblicklichen Momentangeschwindigkeit vm) moduliert sind, werden dazu verwendet, um von externen Quellen erhaltene Signalamplituden in Geschwindigkeits-proportionaler Weise abzutasten. Dies reduziert die Redundanz der Rechenprozesse erheblich (s. auch Fig. 3c). Die solchermaßen durch VTCP-Impulse vm-adaptiv erfassten STQ(d)-Verstreichzeiten werden mit T, die in derselben Weise erfassten STQ(i)-Verstreichzeiten mit T bezeichnet.
Figs. 3b zeigt die Messung von
STQ(d)-Verstreichzeiten mit invarianten ITPC-Impulse mit der Periode
tscan und mit einer konstanten Frequenz fscan. Dies ist der Fall, solange
keine STQ(v)- Parameter erhalten werden, d. h. wenn keine Relativbewegung
zwischen Sensor und Signalquelle auftritt und daher auch keine
Relativgeschwindigkeit (vm) gemessen werden kann.
Figs.
3c zeigt die Messung der
Verstreichzeiten mit modulierten VTCP-Impulsen. Diese Zeitzählimpulse
sind abhängig von der momentanen Relativgeschwindigkeit vm (bzw. vom
erfassten STQ(v)- Parameter), und zwar in einer Weise, dass ihre Periode
(t.v)scan und Frequenz ƒscan proportional zu vm ist. Wenn vm sehr klein
ist oder gegen Null geht, so sinkt die Zählfrequenz ƒscan gleichfalls ab
bis zur Minimalfrequenz fscan (wie in Fig. 3b). Wie in Fig. 2a gezeigt,
wird jeder STQ(v)- Geschwindigkeitsparameter mittels eines zweiten
adäquaten "vorauseilenden" Sensors (bzw. Rezeptors)erfasst. Somit ist vm
schon vor der eigentlichen STQ(d)- und/oder STQ(i)- Verstreichzeitmessung
festgehalten. Es ist daher möglich, ƒscan zur Messung der
Td(1,2...n)-Zeitdaten entsprechend den erfassten STQ(v)- Parametern zu
modulieren, um so die Zahl von t.v- Kalkulationen zu reduzieren sowie
erforderlichen Speicherplatz zu minimieren. Daraus resultiert ein
weitgehend redundanzfreier Analyseprozeß. Obwohl die mit dieser Methode
gezählten Zeitimpulse annähernd kovariant mit der relativ zurückgelegten
Strecke (d) sind, stellen sie dennoch nachweislich modifizierte Zeitdaten
dar und nicht Distanzmaße. Wie auch die Entstehung dieser Daten, so
erfolgt auch die weitere Verarbeitung und Analyse solcher modifizierter
STQ-Verstreichzeiten nach probabilistischen Prinzipien. Die besagten
Zeitdaten werden quasi "subjektiv empfunden". In mechanistischen Systemen
geschieht die annähernd weg-proportionale Modulation der Zeitzählfrequenzen
überwiegend mittels programmierbare Oszillatoren und Timer,
wie aus Fig. 5 ersichtlich ist. Hingegen wird dieser adaptive Prozess (ein
Teil der sogenannten "autonomen Adaption") in komplex strukturierten
biologisch/ chemischen Organismen oft durch proportionale Änderung der
Fortleitungsgeschwindigkeit von Zeitmessimpulsen in neuronalen Fasern
erzielt, wie dies in Fig. 4a, 4b, 4c und 4d gezeigt wird. Autonome
Adaption und adaptive Zeitbasisänderungs-Prozesse der beschriebenen Art
können aber auch anders gestaltet sein. Sie können auf molekularer,
atomarer und subatomarer Ebene existieren. Der Patentanmelder bezeichnet
dieses Prinzip als "zeitliche Auto-Adaption.
Fig.
3d - f zeigen die konzeptionelle
Basis für den Vergleich von aktuell erfasster STQ-Zeitdatensequenzen mit
früher registrierten STQ- Zeitdatensequenzen, sowie ihre Analyse auf
statistischer Grundlage. Die vm-modulierten, in Fig. 3d gezeigten
Zeitdaten Td(n...) mit der Reihe 32-30-22-23-20 (cs= Zählzyklen) werden
Date für Date mit früher registrierten, gleichfalls in vm-modulierter
Weise erfassten Td'-Zeitdaten der Reihe 30-29- 22-24-19 verglichen. Der
Vergleichsprozeß ist genau genommen eine Kovarianzanalyse. Wenn die
Regressionskurven beider Zeitdatenreihen konvergieren, so besteht
Kovarianz. In mechanistischen Systemen kommen zu diesem Zweck
Koinzidenz-Meßsysteme, Komparator-Schaltkreise, Software für statistische
Analysen oder auch "Fuzzy-Logic" zum Einsatz. Die
Wahrscheinlichkeitsdichte-Parameter werden summiert, und sobald der
Gesamtwert innerhalb einer bestimmten Periode eine vorgegebene Schwelle
10 übersteigt, wird ein Signal 11 produziert, welches
anzeigt, dass die Sequenz "erkannt" worden ist. Dieses Signal dient
vorwiegend zur adaptiven Regelung der Aktuatoren in mechanistischen
Systemen (bzw. zur Motorik in Organismen). Weiters zeigt das Signal an,
dass "autonome Adaption" an die registrierten Zeitdatenmuster
stattgefunden hat. Was das motorische Verhalten irgendeines
mechanistischen oder biologischen Mechanismus betrifft, so ist es klar,
dass die Erkennung von Signalfolgen Hand in Hand mit automatischer
Anpassung bzw. autonomer Adaption geht. Dieses Prinzip sei hiermit als
"motorische Auto-Adaption" oder "Auto- Emulation" bezeichnet.
Fig. 3g zeigt diesen
Autoadaptions-Prozess in schematischer und leicht verständlicher Weise.
Eine aktuell erfasste Td- Zeitdatensequenz wird fortgesetzt mit einer
früher erfassten Td'- Zeitdatensequenz verglichen, und besteht annähernde
Übereinstimmung, so passen die Sequenzen zueinander wie Schlüssel und
Schloss. Wie aus den folgenden Beschreibungen ersichtlich, schafft dieser
Vorgang eine Art "Bootstrapping-Prozess" bzw. ein "motorisches
Nacheiferungsbestreben", das eine grundlegende Eigenschaft aller
redundanzfreien autonomen selbstorganisierenden Systeme bzw. eines
Organismus darstellt. Zwar ist die Konvarianzanalyse zweier
Zeitdatenmuster in mechanistisch/elektronischen Systemen relativ
aufwendig (s. dazu Fig. 5); dies gilt jedoch nicht für
molekular/biologische Organismen und andere Einheiten. In solchen
Systemen tritt dieses "Bootstrapping" als "synergetischer Effekt"auf -
annähernd vergleichbar mit dem Rollen einer Anzahl von Billardkugeln in
eine Reihe von Löchern, die ein Muster bilden. (Die Bezeichnung
"Synergetik" wurde übrigens erstmals von H. HAKEN im Jahre 1970
eingeführt). Das erfolgreiche Einlochen wird von Geschwindigkeit und
Richtung bestimmt. Ändern sich diese Parameter, so erfolgt kein
Einlochen. Ein Versuch kann ebenfalls scheitern, wenn sich die Position
der Löcher irgendwie verändern würde, während die Abstoß-Positionen der
Kugeln konstant blieben; selbst dann, wenn ihre Geschwindigkeit und
Richtung kovariant mit den urprünglichen Parametern wäre (und wobei die
Kovarianz die Änderung der Muster nicht entsprechend berücksichtigt). In
ähnlicher Weise prägt eine aktuelle STQ- Zeitdatensequenz, die von einem
autonomen selbstorganisierenden System erfasst wurde, ein bestimmtes
charakteristisches Finger- abdruckmuster, und sooft ein früher
registriertes Referenz-Muster gefunden wird, das isomorph mit einem
aktuell registrierten Muster ist, so resultiert daraus Auto-Adaption und
Auto-Emulation (Nacheiferungsbestreben). Dieses Phänomen wohnt als
teleologisches Ordnungsprinzip allen Lebensformen, Organismen und
elementaren Strukturen inne. Findet sich kein kovariantes referentes
Muster, so kollabiert die auto-adaptive Regulation und das System verhält
sich chaotisch. Der Zustand geht erst dann wieder vom Chaos zur Ordnung
über, wenn aktuell erfasste STQ-Zeitmuster wieder mit früher erfassten
STQ- Zeitmuster zu konvergieren beginnen, die vom Analysator als
"kovariant" befunden werden.
Figs. 4a - d illustrieren ein
Modell der Erfassung und Verarbeitung von STQ(d)- und STQ(v)-
Verstreichzeiten (s. auch Fig. 3a - g) - sowie zur zeitlichen und
motorischen Auto-Adaption - in molekular/biologischem Zusammenhang. Die
grundlegenden Elemente des Modells sind in der Neurophysiologie bereits
von KATZ, GRAY, KELLY, REDMAN, J. ECCLES u. a. beschrieben worden. Die
vorgestellte Erfindung ist aber deshalb von besonderer Neuheit, weil hier
zum ersten Mal zeitliche und motorische Auto-Adaption auf Basis von
STQ-Quanten beschrieben wird. Derartige Systeme bestehen zumeist aus
einer Vielzahl von Neuronen (Nervenzellen). Diese Neuronen kommunizieren
mit Rezeptoren (sensorischen Neuronen), welche die Erfas- sung und
Erkennung der ihrer physikalischen Umgebung ermöglichen. Zusätzlich
kooperieren sie auch mit Effektoren (Muskeln etc.) die als ausführende
Organe für die motorische Aktivi- tät dienen. Der Ausdruck "Rezeptor"
oder "sensorisches Neuron" entspricht dem mechanisti- schen Begriff
"Sensor". Ein "Effektor" ist dasselbe wie ein "Aktuator", wie man ihn aus
der Kybernetik-Literatur kennt. Jedes Neuron besteht aus einer
Zellmembran, die den Zellkern und den Zellinhalt umschließt. Eine
unterschiedliche Anzahl von Fortsätze aus den Neuronen, (Axone,
Dendriten) leitet Information zu Effektoren oder anderen Neuronen weiter.
Die Verbindungsstelle einer dendritischen oder axionalen Endigung mit
einer anderen Zelle wird "Synapse" genannt. Die Neuronen selbst können
als komplexe biomolekulare Sensoren und Zeittaktgeber aufgefasst werden;
die besagten Synapsen als Zeitdatenanalysatoren, welche die aktuell
registrierten Verstreichzeitsequenzen fortgesetzt mit früher
registrierten Verstreich-Zeitmustern vergleichen, welche von den
sensorischen Neuronen (Rezeptoren) produziert und entlang Nervenfasern zu
den Synapsen weitergeleitetet werden. Dort erfolgt eine Art von
"Kovarianz-Analyse", und es werden entsprechende
Wahrscheinlichkeitsdichte- Signale generiert, die an benachbarte
neuronale Systeme oder an Effektoren geleitet
werden.
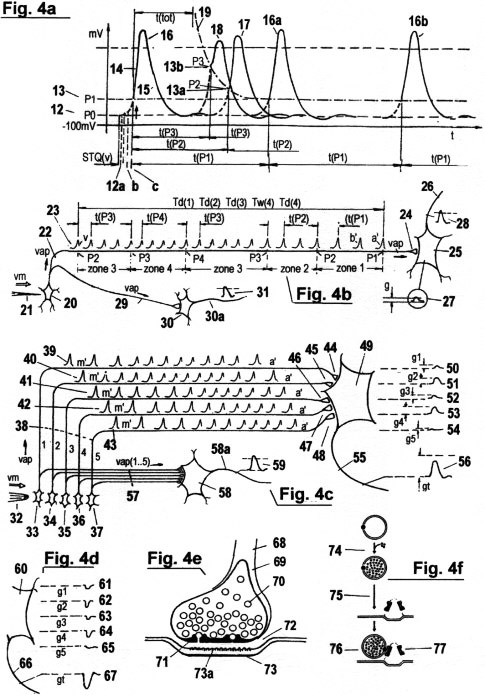
4a zeigt ein sogenanntes "Aktionspotential" AP, das durch
eine abrupte Änderung der Verteilung von Natrium-Ionen und Kalium-
Ionen zwischen dem intrazellulären und extrazellulären Lösungsgemisch
entsteht,
welches wie ein Kondensator arbeitet. Diese Ionen-Konzentrationen
behalten ein bestimmtes Gleichgewicht, solange kein Stimulus an der
Rezeptor-Zelle auftritt.
In diesem Gleichgewichts-Zustand ist ein
konstantes negatives Potential 12 an der Zell-Membran vorhanden,
das "Ruhe-Potential" genannt wird. Sobald ein Rezeptor einen Stimulus aus
einer externen Signalquelle wahrnimmt, fließen Na+ Ionen in die
neuronale Zelle und bewirken, dass sich die Verteilung positiver und
negativer Ionen plötzlich umkehrt, und die Zell-Membrane
"depolarisiert" wird. Je nach Intensivität dieses Rezeptor-Reizes
entstehen nun folgende Effekte:
(a) Wird die Schwelle P1 nicht erreicht, so entsteht ein sogenanntes
"elektrotonisches Potential" EP, das
sich entlang der Zell-Membran (bzw. der axonalen Faser) passiv
fortpflanzt, und in bezug auf
zurückgelegte Zeit und Ort seines Auftretens exponentiell
abnimmt. Die Produzierung eines EP
ist vergleichbar mit dem Anzünden einer leeren Zündschnur.
Das Feuer wird sich ein Stück weit
ausbreiten, dann im weiteren Verlauf schwächer werden
und schließlich verlöschen. EP's entstehen
bei jeder Reizung eines Neurons.
(b) Wird die Schwelle P1
überschritten, so entsteht ein "Aktionspotential" AP (wie in Fig.
4a), das sich entlang der
Zell-Membran (bzw. der axonalen Faser) mit konstanter Amplitude
in selbstregenerierender Weise
fortpflanzt. Die Produzierung eines AP ist vergleichbar mit
dem Auftreten eines Funkens an einer
intakten Zündschnur; das entflammte Zündpulver
erhitzt benachbarte Abschnitte der
Zündschnur soweit, dass dort das Pulver ebenfalls
entflammt usw., wodurch sich die Flamme
entlang der Zündschnur fortpflanzt.
AP's werden zur Quantisierung von STQ(d)- und STQ(v)-Verstreichzeiten verwendet. Sie sind praktisch äquivalent zu jenen Identifikations-Impulsen IP mit den Perioden t(P1), (t(P2), (t(Pn)..., welche in Fig. 3a gezeigt werden. AP's zeigen das Auftreten von Phasen-Übergängen an, von denen STQ(d)-STQ(v)- Verstreichzeiten abgeleitet werden. Zusätzlich bewirken sie indirekt die Aktivierung molekular/biologischer "Zeitmesser" zur Aufnahme solcher Zeiten. AP's stellen jedoch weder deterministische Samplingraten für irgendeine Amplitudenabtastung dar, noch entsprechen sie elektronischen Spannungs/Frequenz-Wandlern. Außerdem ist ihre Amplitude unabhängig von der Reizintensivität am Rezeptor, und sie stellen auch keinerlei Zeittakt-Impulse für die Messung von Verstreichzeiten dar. Hingegen wird die Erfassung solcher STQ-Verstreichzeiten von den Geschwindigkeiten beeinflusst und moduliert, mit denen die Aktionspotentiale sich entlang den Nerven-Fasern (Axonen) und den Membran-Distrikten fortpflanzen. Die zeitmessenden Eigenschaften von AP's werden hier wie folgt detailliert beschrieben: Wenn ein EP - in Antwort zu einem Rezeptor-Reiz - einen bestimmten Schwellwert (P1) 13 überschreitet, dann löst es ein AP 16 aus. Der Amplitudenverlauf eines AP fängt mit dem Aufstrich 14 an und endet mit der Repolarisation 15, beziehungsweise mit der sogenannten "Refraktärphase". Nach dem Ende dieses Vorganges kehrt das Membran-Potential wieder zum Ruhepotential P0 zurück, und die Ionen-Verteilung gelangt wieder ins Gleichgewicht. Nicht jeder Rezeptor-Stimulus erzeugt genügende elektrische Leitfähigkeit, um ein AP zu produzieren. Solange er unter einer Minimalschwelle P1 bleibt, erzeugt er nur das zuvor erwähnte elektrotonische Potential EP. (Für ein besseres Verständnis von Verstreichzeitmessungen in biologisch/chemischen Modellen betrachte man nochmals Fig. 2c und Fig. 3a): Das erste AP, das nach Stimulieren eines Receptors ausgelöst wird, erzeugt zunächst (indirekt) jenen Impuls, der den ersten Zeitmesser zur Aufnahme der ersten STQ(d)- Verstreichzeit aktiviert, wenn die Signalamplitude W das Schwellwert-Potential P1 bei Phasenübergang iTw(1.1) durchbricht. Gleichzeitig stellt es auch einen Identifikations-Impuls IP dar. Das erste AP entspricht quasi dem ersten IP aus einer Reihenfolge von IP 's, die den jeweiligen Status des Schwellenwerts (oder Wahrnehmungbereichs) anzeigen, wo sich die Stimulations- Intensitivität gerade befindet. So lange der Rezeptor-Reiz bestehen bleibt, wiederholt sich ein AP 16a, 16b... in zeitlichen Intervallen, deren Periode von der Höhe des jeweiligen Schwellwertes abhängt, in dem sich der Stimulus gerade befindet. Diese Intervalle entsprechen jenen IP- Perioden t(P1), t(P2)... welche für die serielle Zuordnung und Verarbeitung von STQ- Verstreichzeiten erforderlich sind (s. Fig. 3a). Die AP-Frequenz wird stabilisiert durch die sogenannte "relative Refraktärphase" (Ausfallzeit) nach jedem AP, während derer keine neue Depolarisation möglich ist. Weil sich die Refraktär- Phase in adaptiver Weise proportional zu einer zunehmenden Reiz-Intensität beim Rezeptor verkürzt (z. B. wenn das EP einen höheren Schwellwert P2 bzw. Wahrnehmungs-Bereich 13a erreicht), besteht demnach eine Ähnlichkeit zu einem "programmierbaren bistabilen Multivibrator" wie er in der mechanistischen Elektronik zu finden ist. Der Verlauf dieser Ausfallzeit ("Refraktorität") nach einem AP folgt der strichpunktierten Linie 19.
Fig. 4a zeigt, daß es nach dem Ende einer Repolarisation eine "absolute Refraktoritäts-Periode" t(tot) gibt. Kein neues AP kann während dieser Zeit ausgelöst werden; egal, wie hoch die Reiz- Intensität am Rezeptor steigt. Auf diese Weise ist das "maximale Ausmaß" einer gerade noch wahrnehmbaren Reizintensität programmiert. Wichtig ist die Tatsache, dass sowohl die Dauer der relativen Refraktärphase als auch die Charakteristik der absoluten Refraktoritätsphase auto-adaptiven Gesetzmäßigkeiten unterworfen sind, und sich daher kontinuierlich an neu auftretende Zustandsveränderungen im Organismus anpassen. Das bedeutet, dass auch jene Schwellwerte P0, P1,...., aus denen STQ-Quanten abgeleitet werden, keine absoluten Größen sind, sondern adaptiven Veränderungen unterliegen, wie alle anderen Parameter, insbes. "Zeit", auch. Nun wird erklärt, was nach der Registrierung einer erster STQ(d)-Verstreichzeit bei P1 durch das erste "AP" weiter geschieht: Steigt die Reizintensität (mit einer theoretischen Amplitude W) von der niedrigeren Schwelle P1 zu einer nächsthöheren Schwelle P2 an, dann verursacht das nächste folgende AP indirekt die Registrierung der zweiten STQ(d)- Verstreichzeit, sobald der Phasenübergang durch die nächsthöhere Schwelle P2 erfolgt. Derselbe Vorgang wiederholt sich in Hinsicht auf die Schwellwerte P3, P4 usw. In jedem Fall fungiert das AP gleichzeitig als Identifikations- Impuls IP, wie zu Fig. 3a beschrieben. Es wiederholt sich deshalb in schwellwertabhängigen Perioden solange, als eine Wahrnehmung auf den Rezeptor einwirkt (bzw. solange der Rezeptor etwas "wahrnimmt"). Als Beispiel sei Fig. 3a betrachtet: Solange die Reizintensität in Zone P2 verweilt, kehrt das AP 17, 17a, 17b... in kurzen zeitlichen Perioden wieder. Diese Perioden (oder Intervalle) entsprechen jenen der IP-Identifikations-Impulse mit der Periode t(P2), welche zur seriellen Registrierung der STQ-Verstreichzeiten Td(2) und Tw(2) erforderlich sind. Erreicht die ansteigende Reizintensität den Schwellwert P3 (bzw. die Wahrnehmungsbereichszone 3) 13b, dann wiederholt sich das AP 18a, 18b, 18c... in noch kürzeren Zeitperioden. Dies entspräche jenen IP-Identifikations-Impulsen mit der Periode t(P3) in Fig. 3a , welche indirekt für das serielle Timing der STQ-Verstreichzeiten Td(3) und Tw(3) erforderlich sind. Eine noch größere Reizintensität, zum Beispiel in P4 (oder in Zone 4), würde eine noch kürzere Periode von AP 's erzeugen. Dies entspräche etwa t(P4) in Fig. 3a. Die maximal mögliche AP-Impuls- Frequenz wird jedoch von t(tot) bestimmt. Kürzere Refraktäritäts-Phasen nach der Depolarisation von APs produzieren auch kleinere AP-Amplituden. Diese Eigenschaft vereinfacht die Zuordnung von AP 's zusätzlich. Im folgenden Teil wird die Generierung der eigentlichen Zeitzählimpulse für die STQ- Quantisierung beschrieben. Diese Zeitzählimpulse sind entweder invariable ITPC oder vm- proportionale VTCP wie sie in Fig. 3a gezeigt wurden. Wie erwähnt, sind die Zeitzählimpulse für das Quantisieren von Verstreichzeiten durch jene Geschwindigkeit bestimmt, mit dem sich ein AP entlang eines Axons fortpflanzt. Diese Geschwindigkeit hängt weiters ab vom Ruhepotential und von der Menge des Na+ Ionen-Stroms in den intrazellulären Raum beim Beginn des Depolarisations-Prozesses, sobald die Wahrnehmung an der Rezeptor-Zelle einen elektrischen Strom verursacht, der das extra/intra-zellulare Ionen-Gleichgewicht beeinflusst. Am Beginn der Stimulation eines Rezeptors (dem Ausgangspunkt der Wahrnehmung), fließt nur kapazitiver Strom vom extra-zellularen Raum zur intra-zellularen Flüssigkeit. Dies ruft das besagte "elektrotonische Potential " EP hervor, der sich passiv fortpflanzt. Erst dann, wenn dieses EP die Schwelle P1 überschreitet, wird ein AP produziert, das sich in selbstgenerierender Weise entlang den Membran-Distrikten fortpflanzt. Je mehr kapazitiver Strom noch nach der Depolarisation (bzw. Umladung) des Membran-Kondensators während der Stimulus-Initialisierung verfügbar ist, desto größer ist die Na+-Strömung in den intra-zellularen Raum, und desto mehr EP-Strom kann in noch nicht depolarisierte Gebiete fließen. Auf diese Weise wird das Tempo weiterer Depolarisations-Prozesse in den Nervenfasern, und somit auch die Fortpflanzungsgeschwindigkeit der weiteren AP's, proportional erhöht. Die Umladungszeit des Membran-Kondensators ist also jener Parameter, der die Größe des Ruhepotentials 12 bestimmt. Startet ein Reiz vom niedrigsten Ruhepotential P0 12 aus, so ist der Na+ Einstrom am größten, der Anstieg des EP ist am steilsten und seine elektrotonische Ausbreitung ist am schnellsten. Wird ein AP ausgelöst, dann ist in diesem Fall auch seine Fortleitungsgeschwindigkeit maximal. Aber wenn ein Rezeptor-Stimulus von einem höheren Potential 12a, 12b oder 12c... startet, dann ist der Na+ Einstrom teilweise in- aktiviert, und es nimmt die Steilheit des EP-Anstiegs sowie die Ausbreitungsgeschwindigkeit des elektrotonischen Flusses ab. Deshalb nimmt auch die Fortpflanzungsgeschwindigkeit eines AP ab. Diese besonderen Eigenschaften werden in molekular/biologischen Modellen benutzt, um entweder invariante Zeitzählimpulse ITCP (mit Perioden tscan) oder variable Zeitzählimpulse VTCP (mit Perioden t.vscan) zu produzieren. In letzterem Fall werden die VTCP's entsprechend der relativen Geschwindigkeit vm (via STQ(v)- Parameter) moduliert, und weisen daher entsprechend kürzere Intervalle auf (siehe dazu Fig. 3b und 3c). Das STQ(v)- Quantum wird von der Abweichung des jeweiligen Start-Potentiales in bezug auf das niedrigste Ruhepotential bestimmt, das als "Referenzwert" dient, und wird durch die Dauer der kapazitiven Ladung einer Zell-Membran gemessen, sobald ein Reiz am Rezeptor auftritt. Diese Ladedauer ist umgekehrt proportional zur Geschwindigkeit des Na+ Einstromes durch die Membran-Kanäle in den intra-zellularen Raum. Eine Zellmembran kann als elektrischer Kondensator betrachtet werden, in dem zwei leitende Medien (die intrazelluläre und die extrazelluläre Lösung) durch eine dünne Isolationsschicht, die Membran, voneinander getrennt sind. Beide Medien bestehen aus verschiedenen Arten von Na/K/Cl- Ionen- verteilungen. Je höher die "Stimulus-Dynamik" (siehe unten) ist, welche zuerst das äußere molekulare Medium beeinflusst - das dem Sensor 2 in Abb. 2a entspricht -, und dann das innere molekulare Medium - das dem Sensor 1 entspricht -, desto schneller ist der Na+Einstrom und desto kürzer ist die Ladedauer (welche die Parameter für die Relativgeschwindigkeit vm bestimmt) und desto höher die AP- Fortpflanzungsgeschwindigkeit v(ap) in den benachbarten Membran-Distrikten. Die Signalverläufe an der äußeren und der inneren Seite (bzw.der Membran) entsprechen quasi den Signalamplituden V und W. Die Geschwindigkeit v(ap) generiert also indirekt die beschriebenen invarianten Zeitzählimpulse ITCP oder die variablen vm-proportionalen Zeitzählimpulse VTCP. Die besagten variablen VTCP-Impulse sind selbst-adaptiv modulierte Zeitimpulse mit Korrelation zur relativ zurückgelegten Distanz. Wie im folgenden Abschnitt erklärt wird, existiert (im Gegensatz zu traditionellem physikalischen Verständnis) keine invariante Zeit - nur "wahrgenommene Zeit" ist tatsächlich existent. Von wesentlicher Wichtigkeit ist auch der Unterschied zwischen "Reiz-Intensität", deren Maß von der AP-Frequenz und somit durch die Refraktär-Phase bestimmt wird, und der "Reiz-Dynamik", deren Maß durch die Ladedauer der kapazitiven Zell-Membran - und deshalb von der Geschwindigkeit des Na+ Zustromes - definiert wird. "Reiz- Dynamik" ist nicht gleichzusetzen mit "Reiz-Intensitätszunahme". Es ist ein Maß für die räumlich/zeitliche Abweichung der relativen Position des Rezeptors in bezug auf die Position der Stimulus-Quelle, und deshalb für die relative Geschwindigkeit vm. " Reiz- Intensität" steht für jene Signalamplituden, aus denen vm-adaptive STQ(d)-Verstreichzeiten Td(1,2,3..) abgeleitet werden, während "Reiz-Dynamik" durch die erfassten STQ(v)- Parametern definiert wird.
Fig. 4b und Fig. 4c zeigen die Analyse von STQ-Verstreichzeiten in einem molekular/ biologischen Modell in leicht verständlicher Weise. Die Ergebnisse der Analyse werden dazu benutzt, um redundanzfreie auto-adaptive Muster-Erkennung sowie autonome Regelungs- und Selbstorganisations-Prozesse zu generieren. Im gezeigten Beispiel sollte ein Organismus in die Lage versetzt werden, bestimmte Arten von Fremdkörper, die auf eine Hautpartie drücken, voneinander zu unterscheiden. Er soll mit einem schnellen Muskel-Reflex antworten, wenn er einen Nadelstich als solchen erkennt. Hingegen sollte er den Reiz ignorieren, wenn es sich um einen stumpfen Gegenstand handelt. Dazu ist eine fortgesetzte vm-adaptive Erfassung von STQ(d)-Verstreichzeiten mittels VTCP-Impulse notwendig. Die Frequenz dieser Zeitzählimpulse wird entsprechend den STQ(v)- Parametern aus der Reiz-Dynamik (vm) moduliert. Diese STQ(v)- Parameter werden zur Registrierung der STQ(d)-Verstreichzeiten Td(1,2,3...) aus der Signal-Amplitude der gegenwärtigen Reiz-Intensität benötigt. Der Unterschied zwischen "Reizintensität" und "Reizdynamik" ist in diesem Beispiel leicht zu verstehen. Ein Stimulus kann verschiedene Intensität auch dann aufweisen, wenn keine zeitlich/räumliche Änderung zwischen Signalquelle und Rezeptor stattfindet. Eine Nadel in der Haut kann auch bei unveränderter Position ein unterschiedliches Empfindungsmuster bewirken, beispielsweise wenn sie erwärmt wird. Dieses Empfindungsmuster wird von der Signalamplitude und somit von der AP-Frequenz und den STQ(d)-Quanten bestimmt. Befindet sich die Nadel in einer invarianten Position, so ist die AP-Fortleitungsgeschwindigkeit konstant, da auch die Membranladezeit konstant ist. Während des Stiches in die Haut ereignet sich ein "dynamischer Reiz", und die STQ(d)-Quantisierung der Signalamplitude geschieht in Abhängigkeit vom Verlauf der Einstichgeschwindigkeit vm. Man beachte, daß während dieses dynamischen Prozesses immer 2 zeitlich verschobene Signal-Amplituden (an der äußeren und der inneren Membranseite) existieren. Die beschriebenen STQ(v)-Parameter leiten von ihnen her. Dem- entsprechend werden die Geschwindigkeiten der AP- Fortpflanzung und die erworbenen STQ(d)- Zeitmuster angepasst ("zeitliche Auto- Adaption"). Diese entsprechend zu vm adaptiv gemessenen STQ(d)-Zeitmuster Td(1,2,3...) werden ständig mit vorausgehend gemessenen und gespeicherten STQ(d)- Zeitmustern Td'(1,2,3...) verglichen und zusammen ausgewertet. Dieser Zeitvergleichs-Prozess geschieht fortgesetzt in den sogenannten "Synapsen", welche die Verbindungsstellen zu axionalen Endigungen anderer Neuronen bilden. Die Wahrscheinlichkeitdichte-Werte, die in den Synapsen ermittelt werden, und die für eine Konvergenz beider Regressions-Kurven stehen, werden zur weiteren Verarbeitung zu peripheren neuronalen Systemen, oder auch an Muskelfasern weitergeleitet, um motorischen Reflex hervorzurufen.
Fig. 4b zeigt die vm- abhängige
Fortpflanzung eines AP von einem Sensor-Neuron (Rezeptor)
20 entlang eines Neuronenfortsatzes (Axon) zu einer Synapse,
wo ein Vergleich mit erworbenen Zeit-Sequenzen mittels molekularer
"Kovarianzanalyse" stattfindet. Dieser Rezeptor funktioniert wie
ein "Druck-Sensor". Wenn eine Nadel 21 mit einer bestimmten Dynamik
auf die äußere Schicht der Zell-Membran trifft, dann verursacht
diese Stimulation das Auslösen von AP's 23, wie in Abb. 4a
beschrieben. Die AP's pflanzen sich im Axon 22 mit einer STQ(v)-
abhängigen Geschwindigkeit vap fort. Ihre Reihenfolge (a'....v ')
entspricht jenem Signal- amplitudenverlauf, der vom Stich
hervorgebracht wird. Die Reihenfolge fängt mit dem Phasendurchgang beim ersten Schwellwert P1 an, geht weiter über P2, P3, P4
(an deren Stelle das Reizmaximum erreicht wird), und kommt
schließlich zu den Phasendurchgängen P3 und P2. Die
Intensitätsbereiche der Reizwahrnehmung sind mit Z1, Z2, Z3 und Z4
bezeichnet. Die Perioden t(P1), t(P2), t(P3), t(P4).... und die
Magnituden der AP's dienen zur Identifikation der jeweiligen
Schwelle, wo die Reizintensität sich gegenwärtig befindet. Ihre
zeitliche Reihenfolge ist deshalb eine Art von "Code". AP's sind
keine Zeitzählimpulse. Neben ihrer Code- Funktion sind sie aber auch
(indirekt) Startimpulse und Stopimpulse für die Registrierung von
STQ(d)- Verstreichzeiten. Die eigentliche vm-abhängige Messung der
STQ-Verstreichzeiten Td(1), Td(2), Td(3), Tw(4) und Td(4)..(s. Abb.
2c), sowie ihr Vergleich mit vorausgehend erfassten
Verstreichzeiten findet in der Synapse 24 statt.
Am
präsynaptischen Ende des Axons laufen die AP 's 23 mit variablen
Geschwindigkeiten vm(n ...) ein, die in Relation zur Dynamik des
Nadel-Stiches sowie zu den gemessenen STQ(v)- Parametern stehen.
Diese variablen Einstrom-Geschwindigkeiten an den Synapsen sind der
Schlüssel zum Produzieren jener adaptiven Zeitzählimpulse VTCP
(s.Fig. 3c) mit der vm- modulierten Frequenz ƒscan. Die Synapse ist
durch den "synaptischen Spalt" von der postsynaptischen Membran
getrennt; diese wiederum ist mit anderen Neuronen verbunden, z. B.
mit einem sogenannten "Motoneuron" 25. Ein solches Neuron
produziert ein sogenanntes "exzitatorisches postsynaptisches
Potential" (ESPS) 27, das etwa proportional zur Wahrscheinlichkeit
g einer Konvergenz ist. Wenn dieses EPSP (bzw. die
Wahrscheinlichkeitsdichte g) einen bestimmten Schwellwert
übersteigt, dann wird erneut ein Aktionspotential AP 28
ausgelöst.Dieses AP wird über Motoaxon 26 zu einer sogenannten
"neuromuskulären Endplatte" weitergeleitet, die dort den
Muskelreflex verursacht. Die einlaufenden AP- Sequenzen 23 bewirken
die Freisetzung bestimmter Quanten von molekularem Transmitter-
stoff aus ihren Depots - winzigen kugelförmigen Strukturen in der
Synapse, "Vesikel" genannt. Im Prinzip stellt eine Synapse einen
komplexen programmierbaren Zeitdatenprozessor und -Analysator dar,
der immer dann den Inhalt jeweils eines Vesikels in den
präsynaptischen Spalt entleert, wenn das Wiederauftreten
irgendeiner früher registrierten synaptischen Struktur innerhalb
einer neu registrierten Schlüssel-Struktur bestätigt wird. Die
synaptischen Struktur innerhalb einer neu registrierten
Schlüssel-Struktur Strukturen und Vesikel- Bewegungen werden von der
Dynamik v(ap) des AP-Ioneneinstroms sowie seiner Frequenz erzeugt.
AP-Einstromgeschwindigkeiten v(ap) entsprechen den STQ(v)-
Parametern, die AP-Frequenzen den STQ(d)-Verstreichzeiten. Die
Transmitter-Substanz wird durch die Synapse reabsorbiert und
wiederverwertet, sodaß der Zyklus nicht unterbrochen wird. Hier
eine ausführliche Beschreibung von Fig. 4b unter Bezugnahme auf Fig.
4e und Fig. 4f: Der Ionen-Einstrom vom ersten einlaufenden AP
23
(a') aktiviert zunächst die Ansammlung von ACh-Transmittermolekülen
in den kugelförmigen Strukturen (Vesikeln). Die Dauer dieser
ACh-Ansammlung hängt ab von der Dynamik (= Geschwindigkeit vap) des
AP-Ionen- Einstromes an der präsynaptischen Endigung und somit auch
der Reiz-Dynamik (= vm) am Rezeptor 20. Die Freisetzung der
Moleküle in den synaptischen Spalt erfolgt in Form von "Paketen".
Jedes nacheinander einlaufende AP - bezeichnet mit b', c'..-
bewirkt eine erneute Ansammlung von Neurotransmitterstoffen in
Vesikeln, die sich nach erfolgter Auffüllung in Richtung des
synaptischen Spalts bewegen. Alles Folgende hat
Verstreichzeit-messende und -analysierende Eigenschaften: Die Dauer
der Auffüllung mit Transmittersubstanz T(t), die Geschwindigkeiten
v(t), mit dem sich die Vesikel in Richtung des synaptischen Spalts
weiterbewegen, ihre Einwirkung auf das synaptische Gitter am Spalt
und ihre Positionen, die Öffnungsdauer der Poren usw. Durch diese
AP-Einwirkung auf die synaptischen Strukturen werden nicht nur die
eigentlichen Zeitzählfrequenzen ƒscan generiert (zur vm-abhängigen
Messung jener in Abb. 2c beschriebenen STQ(d)-Versteichzeiten),
sondern es werden auch Zeitmuster gespeichert und analysiert. Wenn
von der Synapse das Muster eines aktuellen zeitlichen Ablaufes
erkannt wird, das mit einer bestehenden gespeicherten Struktur
übereinstimmt, dann öffnet sich eine Pore beim synaptischen
Gitter, und der ganze molekulare Inhalt eines Vesikels wird in die
subsynaptischen Spalt freigesetzt. Die freigesetzten Transmitter-
Moleküle (zumeist ACh) binden sich an der anderen Seite des Spalts
an spezifische Rezeptormoleküle auf der subsynaptischen Membran des
angekoppelten Neurons. Dadurch wird dort ein postsynaptisches
Potential (EPSP) hervorgerufen, das sich dann an andere Synapsen,
Dendriten oder an eine sogenannte "neuromuskuläre Endplatte"
fortpflanzt. Überschreitet das EPSP eine bestimmte Amplitude, dann
löst es ein Aktionspotential (AP) der bereits beschriebenen Art
aus, das dann z.B. einen Muskel-Reflex hervorruft. Erreicht es
diesen Schwellwert nicht, so wird es ähnlich weitergeleitet wie ein
EP (also in elektrotonischer Weise); ein AP wird in diesem Fall
nicht produziert. Von besonderer Bedeutung ist die addierende
Eigenschaft der subsynaptischen Membran. Diese Eigenschaft - auch
als "zeitliche Bahnung" bezeichnet - resultiert aus der Summierung
der erzeugten EPSP's, wenn diese in kurzer Folge innerhalb
bestimmter Zeitfenster eintreffen. Jede Freisetzung von
Transmittermoleküle in den synaptischen Spalt zeigt das Auftreten
einer erhöhten Wahrscheinlichkeits-Dichte während des Vergleiches
aktueller vm-proportional erfaßter STQ-Verstreichzeitmuster mit
früheren vm-proportional erfaßten und gespeicherten Zeitmustern an.
Erhöhte Wahrscheinlichkeits-Dichte verursacht eine höhere
Häufigkeit von Transmittersubstanz- Freisetzung und demnach höhere
Summenwerte von EPSP's, was wiederum als Folge eine signifikant
höhere Rate von postsynaptischen Aktionspotentialen (AP) produziert.
Demnach ist ein postsynaptische AP ein Bestätigungssignal, das
anzeigt, dass Isomorphismus zwischen einem früheren und einem
aktuell registrierten Zeitdatenmuster erkannt worden ist. Auf der
Basis dieses Zeitmuster-Vergleiches wird somit jener Gegenstand,
der die Wahrnehmung bei der Rezeptor-Zelle verursacht hatte, als
"Nadel" identifiziert; und der Befehl: "Muskelreflex auslösen" wird
an die korrespondierenden Muskelfasern weitergeleitet.
Parallele komplexere Erkennungsprozesse laufen über das zentrale
Nervensystem ZNS (dem Gehirn) ab. Vom beschriebenen
druckempfindlichen Hautrezeptor-Neuron 20 gelangt eine weitere
axonale Verzweigung 29 über eine Synapse 30 zu einem "ZNS-Neuron".
Im Unterschied zum "Motoneuron", das direkt die motorische
Aktivität des Organismus steuert, dient ein ZNS-Neuron der
bewussten Erkennung eines rezeptorischen Reizverlaufes. Ein AP
31,
das an der postsynaptischen Zellmembran 30 produziert wird, kann
auch entlang Dendriten in ein Axon 30a münden, und sich zu mehreren
anderen ZNS-Neuronen verzweigen; es kann aber auch indirekt über
ZNS-Neuronen zu einem Motoneuron und dann zu einer "neuromusculären
Endplatte" gelangen.
Die Parameter, welche die
Registrierung von STQ-Zeitquanten in den Synapsen 25 und
30
kontrollieren, können sich durch die Verschiedenheit der
synaptischen Strukturen unterscheiden. (Tatsächlich werden ja
diese Strukturen von fortgesetzten "Lern"-Prozessen erzeugt). Dies
erklärt, warum es möglich ist, dass ein Nadelstich zwar im Gehirn
als solcher registriert wird, aber keinen Muskel-Reflex
hervorruft; oder aber ein schneller Muskel-Reflex entstehen kann,
dessen Ursache durch das Gehirn kaum wahrgenommen wird. Der eine
Fall zeigt einen bewussten Reflex, der andere Fall einen
instinktiver Reflex. Letzterer tritt auf, wenn die ZNS-Synapse
30
nicht genügend isomorphe Strukturen findet (im Gegensatz zur
Synapse 25), daher keine ausreichend häufige Freisetzung von
Transmittermolekülen erfolgt, und infolgedessen auch kein
postsynaptisches AP 31 und keine bewusste Erkennung des
wahrgenommenen Reizes stattfinden kann.Viele Funktionen des
zentralen Nervensystemes können so auf monistischer Grundlage
erklärt werden; sogar Phänomene wie "Bewusstsein" und
"Unterbewusstsein". Im allgemeinen sind auto-adaptive Prozesse in
Organismen vielseitig vernetzt und deshalb äußerst komplex. Um
fähig zu sein, auf der Haut einen Nadelstich vom Drücken mit einem
stumpfen Radierstift zu unterscheiden, sind wesentlich mehr
Zeitmuster notwendig; auch müssen mehr Rezeptoren und Synapsen in
den Erkennungsprozess einbezogen werden.
Fig. 4c zeigt
einen Prozess, bei dem das leichte Drücken mit einem stumpfen
Gegenstand (z.B. einem konischen Radiergummi auf einem Stift)
erkannt wird - wo aber daraus kein Muskelreflex resultiert. Der
stumpfe Gegenstand 32 presst mit einer bestimmten relativen
Geschwindigkeit vm auf eine Reihe von Rezeptoren in den neuronalen
Hautzellen 33, 34, 35,
36 und 37. Aus der Stimulierung der
einzelnen benachbarten Rezeptoren (s. auch Fig. 4b) resultieren
verschiedene Sequenzen von AP's 39, 40,
41, 42 und 43. Diese
Aktionspotentiale pflanzen sich entlang den kollateralen Axonen
38
mit variablen Perioden t(P1,2,3.....) und Geschwindigkeiten
vap(1...5) fort, die einerseits von der gegenwärtigen
Reizintensität, anderseits von der jeweiligen Reizdynamik abhängig
sind. Da jeder Rezeptor-Reiz ein anderes Muster aus STQ(v) und
STQ(d)- Quanten erzeugt, ergeben sich für jedes Axon verschiedene
AP-Sequenzen a'....m'. Alle einzelnen Sequenzen zusammen stellen
jenes Muster aus STQ- Verstreichzeiten dar, das für den Druck mit
dem Radierer auf der Haut charakteristisch ist. Diese variablen
AP-Ionenströme gelangen zu den Synapsen 44, 45,
46, 47 und 48,
welche über den synaptischen Spalt mit dem Motoneuron 49 verbunden
sind. Sobald das aktuell erfasste Zeitdatenmuster eine Ähnlichkeit
zu einem früher erfassten Zeitdatenmuster aufweist, setzt jede
einzelne Synapse den Inhalt eines Vesikels in den subsynaptischen
Spalt frei. Gleichzeitig produziert diese Freisetzung ein ESPS an
der subsynaptischen Membran des Neurons. Diese Potentiale sind
zumeist unterschwellig. Der nötige Schwellwert für die Auslösung
eines AP wird nur dann erreicht, wenn mehrere EPSP's summiert
werden. Dies geschieht nur dann, wenn eine sogenannte "zeitliche
Bahnung" solcher Potentiale stattfindet, wie im Abschnitt zuvor
beschriebenen wurde.
Im gezeigten Modell wirken sich diese
addierenden Eigenschaften auf die einzelnen ESPS's 50,
51, 52, 53
und 54 aus. Diese Potentiale entsprechen den rezeptor-spezifischen
Wahrscheinlichkeitsdichte-Parametern g1, g2. g3, g4 und g5, die für
den jeweilige Grad an Isomorphität von Zeitmustern stehen.
Gleichzeitige Transmitterfreisetzung in mehreren Synapsen (z.B. in
45 und 47) bewirkt die Addierung der einzelnen EPSP's zu einem
Gesamt- Potential 56, das der Summe der einzelnen
Wahrscheinlichkeitsdichten (G) = (g1+ g3) entspricht. Diese
Eigenschaft der Neuronen, räumlich getrennte unterschwellige EPSP 's
zu summieren, wenn gleichzeitige Transmitter-Freisetzung bei einer
Anzahl paralleler Synapsen auf der selben subsynaptischen Membran
auftritt, bezeichnet man als "räumliche Bahnung". Im beschriebenen
Modellfall erreicht das summierte EPSP 56 aber nicht die bezeichnete
Schwelle (gt) und es wird daher kein AP ausgelöst. Statt dessen
pflanzt es sich im subsynaptischen Membrandistrikt 49 des Neurons
bzw. im nachfolgenden Motoaxon 55 als passives elektrotonisches
Potential (EP) fort. Ein solches EP schwächt sich (zum Unterschied
von einem selbst-regenerierenden aktiven AP) nach wenigen
Millimetern im Axon soweit ab, dass es keinerlei aktivierenden
Einfluss auf die neuromuskuläre Endplatte hat, und somit auch
keinen aktivierenden Einfluss auf den Muskel. Die Reizung der Haut
und das Anpressen mit dem Radierstift reicht also nicht aus, um
einen Muskelreflex hervorzurufen. Anders wäre es der Fall, würde
der Radiergummi abbrechen und der leere Stift mit voller Wucht auf
die Haut- Rezeptoren treffen. In diesem Fall würde eine
Transmitterfreisetzung gleichzeitig in allen fünf Synapsen 50,
51,
52, 53 und 54 ausgelöst, weil die erfassten STQ-Zeitmuster
Td(1,2,3....) mit sehr hoher Wahrscheinlichkeit ähnlich mit jenen
in den synaptischen Strukturen gespeicherten STQ-Zeitmustern
Td'(1,2,3..) wären, die für das Ereignis "Nadelstich" signifikant
sind. Die EPSP 's würden wegen der Eigenschaft ihrer zeitlichen und
räumlichen "Bahnung" zu einem überschwelligen EPSP 56 addiert
werden und ein postsynaptisches AP
hervorrufen, das sich im Motoaxon 55 in einer
selbstregenerierenden Weise (ohne zeitliche und räumliche
Abschwächung) bis zum Muskel fortpflanzen würde um einen
Muskel-Reflex zu produzieren. Wie in Fig. 4b, so kommt es auch im
vorliegenden Beispiel zu einem parallel verlaufenden
Erkennungsprozess, der im zentralen Nervensystem (ZNS) stattfindet.
Von den Hautrezeptor-Zellen 33, 34,
35, 36 und 37 gelangen
kollaterale axonale Verzweigungen 57 zu ZNS- Synapsen, die an andere
Neuronen 58 gekoppelt sind. Solche Verzweigungen werden als
"Divergenzen" bezeichnet. Die Aufsplitterung der Axone in
kollaterale Verzweigungen zu verschiedenen neuronalen
ZNS-Distrikten, und die zeitliche und räumliche Kombination vieler
postsynaptischer EPSP's ermöglicht die bewusste Erkennung komplexer
Wahrnehmungen im Gehirn (z.B. die Tatsache, dass ein Radierstift
auf die Haut drückt). Da diese Erkennung unabhängig von der
Auslösung eines etwaigen Muskelreflexes stattzufinden hat, muss die
Summe einzelner ESPS's im ZNS auf jeden Fall überschwellig sein.
Ansonsten könnte kein postsynaptisches AP - und somit auch kein
Bestätigungssignal produziert werden.
Als wesentliche
Vorbedingung dazu ist notwendig, daß auto-adaptive Prozesse
vorangingen, wodurch bestimmte präsynaptische und subsynaptische
STQ-Zeitstrukturen in den parallelen Synapsen 58 geprägt wurden.
Diese Strukturen enthalten Information (Zeitsequenzen,
Zeitmuster..), die für ähnliche Sinnes-Erfahrungen stehen (z. B. in
die Haut eindringende Gegenstände, darunter auch der besagte
Radierstift). Offensichtlich hat die Schwelle für das Verursachen
eines AP's in der postsynaptischen Membran-Struktur der ZNS-Neuronen
58, (und deshalb auch im Gehirn) niedriger zu sein als beim vorher
beschriebenen Motoneuron 49. Daher muss auch die Summe dieser
ESPS's größer sein als diejenige der ESPS's g1, g2, g3, g4 und g5.
Isomorphitäten von STQ-Zeitdatenreihen in den ZNS-Synapsen des
Gehirns haben demnach ausgeprägter zu sein als jene in den Synapsen
von Motoneuronen, die nur für Muskelreflexe zuständig sind. Die
Strukturen von ZNS-Synapsen müssen Informationen besser
unterscheiden können, und somit subtiler beschaffen sein. Das
Auftreten eines sub-synaptischen AP stellt eine Bestätigung für die
Tatsache dar, dass ein aktuell erfasstes Td(1,2,3...)- Zeitmuster
in virtueller Weise isomorph ist mit einem früher registrierten
Td'(1,2,3...)- Referenzzeitmuster, das z.B. aus einer älteren
Sinneserfahrung mit einem an irgendeine Hautstelle drückenden
Radierstift stammt. Hat eine solche frühere Erfahrung nicht
stattgefunden, so hat das Bewusstsein keine physikalische Grundlage
zur Erkennung, da die Zeitmuster-Vergleichsbasis fehlt. In einem
solchen Fall hätte also zunächst ein Lernprozess vorauszugehen.
Meistens werden dazu auch andere, aus verschiedenen Rezeptorreiz-
Ereignissen hergeleitete Empfindungserfahrungen visueller oder
akustischer Art usw. mit der erwähnten Druckempfindungs-Erfahrung
koordiniert.
Dies erklärt, warum ZNS-Strukturen äußerst
umfangreich vernetzt sind. ZNS-Neurone, aber auch Motoneurone,
besitzen bis zu 5000 angekoppelte Synapsen, die in verschiedenster
Weise mit Rezeptor- Neuronen und axonalen Verzeigungen verknüpft
sind. Es existieren komplexe Zeitdaten-Muster für niedrigere und
höhere Aufgabengebiete, die in hierarchischer Weise strukturiert
sind. Einfache Td(1,2,3...) und Td'(1,2,3...)- Analyse-Operationen
wurden bereits beschrieben. Kreislauf, Atmung, Koordination von
Muskelsystemen, Wachstum, Sehen, Hören, Sprechen, Riechen usw.
erfordern eine enorme Anzahl synaptisch registrierter "Landschaften"
von STQ-Zeitmustern im Organismus, die von vielen
Rezeptoren produziert wurden; und welche fortgesetzt auf
Isomorphität mit aktuell aufgenommenen STQ-Zeitmustern analysiert
werden müssen. Dementsprechend ereignet sich zeitliche und
motorische Auto-Adaption in tieferen und höheren Hierarchien und auf
verschiedenen Niveaus.
Fig. 4d zeigt das Gegenstück
zu einem EPSP (erregendes postsynaptisches Potential); das
"inhibitorische postsynaptische Potential", oder IPSP. Wie aus der
Zeichnung zu sehen ist, sind die IPSP- Potentiale 61,
62, 63, 64
und 65 an der subsynaptischen Membran 60 im Unterschied zu besagten
EPSP's negativ. IPSP's werden von einem erheblichen Teil der
Synapsen produziert, um präsynaptische Hemmung statt Aktivierung
hervorzurufen. Das Beispiel zeigt hier ein sich vom Motoaxon
66 zu
einer neuomuskulären Endplatte (beziehungsweise Muskelfaser)
fortpflanzendes Summen-IPSP 67, welches verhindert, dass dieser
Muskel aktiviert werden kann - selbst dann nicht, wenn ein
überschwelliges EPSP zur gleichen Zeit über ein paralleles Motoaxon
die gleiche Muskelfaser erreichen würde.
Positive
EPSP's-Ionenströme und negative IPSP's-Ionenströme heben sich
gegenseitig auf. Die hauptsächliche Funktion von IPSP's besteht
darin, koordinierte und homogene Zustands- veränderungen im
Organismus, z.B. exaktes Timing von Bewegungsvorgängen zu
ermöglichen. Um beispielsweise einen gleichmäßigen Armschwung
durchzuführen, ist es erforderlich, den Musculus biceps, der den
Ellenbogen beugt, mittels EPSP's zu aktivieren; gleichzeitig aber
den antagonistischen Musculus triceps (der den Ellenbogen streckt),
mittels IPSP's zu hemmen. Antagonistische Muskel müssen über
sogenannte "antagonistische Motoneuronen" gehemmt werden, während
der andere Muskel über "homonyme Motoneuronen" aktiviert wird. Der
komplexe Synergismus von erregenden EPSP-Synapsen und hemmenden
IPSP-Synapsen wirkt wie ein Feedback-System (Regelkreis) und
ermöglicht optimales Timing und Effizienz im Organismus. Man kann
diesen Prozess mit einem Servo-Antrieb oder einer Servo- Lenkung
vergleichen, welche eine korrekte koordinierte Ausführung
momentaner Bewegungsphasen durch datenunterstützte Operation und
Regelung ermöglicht. Wenn Daten ausfallen, bricht der Servo-Kreis
zusammen. Störungen in einem molekular-biologischen Regelkreis, der
durch STQ-Zeitdatenstrukturen gestützt wird, führen zu tetanischen
Zuckungen, willkürlichen Kontraktionen, chaotischen Krämpfen
usw.
Vom kybernetischen Standpunkt betrachtet, erzeugt jede
erregende Synapse einen "motorischen Impuls" (EPSP), jede hemmende
Synapse einen "Bremsimpuls" (IPSP). Das stetige Abstimmen der
komplizierten Regelkreise, und die Balance, die aus dem
fortgesetzten Vergleich von früheren sensorischen Erfahrungen (den
gespeicherten referenten Zeitmustern) mit aktuellen sensorischen
Erfahrungen (den aktuell erfaßten Zeitdatenmustern) resultiert,
erzeugt ein "perfektes Timing" im Organismus.
Fig. 4e
zeigt die prinzipielle Gestaltung einer Synapse. Das Axon 68 mündet
in die präsynaptische Endigung 69, die auch "Bouton" genannt wird.
Die seriell einlaufenden AP's verursachen die Auffüllung der
Vesikel mit Neurotransmitter-Mulekülen. Ist der Auffüllprozess
beendet, beginnen sich die Vesikel in Richtung präsynaptisches
Vesikelgitter 71 zu bewegen. Wenn ein gegenwärtig erworbenes
Zeitmuster annähernd isomorph einem bestehenden Zeitmuster ist, (s.
auch Fig. 4b), dann öffnet sich bei einer Andockstelle am
synaptischen Gitter ein kleiner Kanal, der den ganzen Inhalt des
Vesikels in den schmalen synaptischen Spalt 72 freisetzt. Dieser
Vorgang wird auch als "Exozytose" bezeichnet. An der subsynaptischen
neuronalen Membran 73 befinden sich spezifische molekulare
Rezeptoren 73a, an die sich die freigesetzten Transmittermoleküle
heften. Für eine bestimmte Dauer wird eine eine Pore geöffnet,
durch welche die Transmitter-Substanz diffundiert. Die Leitfähigkeit
der postsynaptischen Membran erhöht sich und das besagte EPSP (die
darauffolgende postsynaptische Depolarisation) wird ausgelöst. Die
Öffnungs-Dauer der Poren und die Erkennung komplementärer
Rezeptoren durch die Moleküle wird gleichfalls von auto-adaptiven
Prozessen und Evaluationen von STQ-Zeitmusterstrukturen bestimmt.
Allerdings stellen diese Prozesse Subphänomene von niedrigerer
Stufe im Vergleich zu synaptischen Phänomenen dar.
Strukturen
zur zeitlichen und motorischen Auto-Adaption, die auf der
Quantisierung von STQ-Verstreichzeiten beruhen, existieren auch auf
molekularem und atomarem Niveau.
Fig. 4f zeigt das Auffüllen eines
Vesikels 70 mit Transmitterstoffen und seine darauf folgende
Bewegung zu einem präsynaptischen Andockvorsprung am Vesikelgitter
71. Der Beginn der Auffüllung 74 ist mit der Aktivierung einer
Stoppuhr vergleichbar. Das Tempo v(t) der Vesikel- Auffüllung ist
proportional zur Dynamik des AP-Ioneneinstroms vap in die Synapse.
Die Perioden der Auffüllungen T(t...) richten sich nach den
Perioden t(P1,P2...) der eintreffenden AP's; somit stellen diese
Zeiten quasi vm-adaptiv quantisierte STQ(d)- Verstreichzeiten
Td(1,2,3...) usw. dar. Die Vesikel- Bewegungsrichtung ist mit
75
bezeichnet. Wenn das aktuelle Tempo v(t), die Dauer der
Vesikel-Auffüllung T(t), die Transmitter-Menge, die aktuelle
Vesikel-Bewegung und andere aktuelle signifikante STQ-Parameter
korrelative Eigenschaften zu bestehenden synaptischen STQ-Strukturen
haben, so heftet sich ein aufgefüll- tes Vesikel an einen
Andockvorsprung 77 am Gitter. Ca++ Ionen strömen in die Synapse ein,
eine Pore am kristallinen Vesikelgitter öffnet sich, und der
gesamte molekulare Transmitterinhalt entleert sich in den
synaptischen Spalt 72. An der postsynaptischen Membran der
Zielzelle fusionieren sich diese Moleküle mit den spezifischen
Rezeptor-Molekülen. Diese Rezeptoren haben
Verifizierungs-Aufgaben. Sie verhindern, dass fremde
Transmitterstoffe (die aus anderen Synapsen stammen) an diesem
Neuron falsche ESPS's auslösen könnten. Zur Vervollständigung
dieses Kapitels werden die Beschreibungen zu Fig. 4a, 4b, 4e
und 4f in Kontext zu den STQ-Konfigurationen von Fig. 3c - 3g gebracht.
Als Argumentationsbeispiel wird wieder angenommen, dass ein Nadelstich auf eine
Rezeptor-Zelle trifft (s. auch Fig. 4b): Jene IP-Sequenzen, die in
Fig. 3a gezeigt werden, entsprechen den AP's 23, die durch das
Stimulieren einer Rezeptorzelle 20 mit einer Nadel
21 produziert
werden. Ihre Perioden t(P1), t(P2) ... dienen der Zuordnung der
jeweiligen Zone der Reizintensität (P1, P2...) oder der
Wahrnehmungsintensität (Z1, Z2..). Jedes in einer Synapse 69
einlaufende AP löst eine von seiner axonalen
Fortleitungsgeschwindigkeit (vap)-abhängige adaptive Quantisierung
von STQ(d)-Verstreichzeiten aus. Die Verstreichzeitnahme mit
modulierter Zeitbasis wird ausgelöst, sobald eine
Vesikel-Auffüllung beginnt. Eine beendete Auffüllung bedeutet
"Verstreichzeitnahme gestoppt; STQ(d)-Quantum registriert". Diese
solcherweise registrierten Verstreichzeiten Td(1), Td(2), Td(3),
Td(4)...usw. generieren die signifikanten synaptischen Strukturen.
Invariante Zeitzählimpulse ITCP (s. Fig. 3b) mit der Frequenz fscan
entsprechen einer konstanten axonalen AP-Fortpflanzung mit der
Geschwindigkeit vap, wenn kein dynamischer Stimulus der besagten
Hautrezeptor-Zelle gegeben ist; (z.B. wenn eine Nadel in einer
fixen Position verharrt und eine konstante Reizintensität erzeugt).
In diesem Fall erfühlt die Rezeptor-Membran keine relative
Geschwindigkeit vm, die AP 's pflanzen sich mit fixem vap entlang
dem Axon 22 fort; und die Synapse quantisiert die
STQ(d)-Verstreichzeiten mit invarianter Zeitzählfrequenz
fscan.
Jene Zeitzählimpulse VTCP (s. Fig. 3c) mit der
variablen Frequenz ƒscan kommen dann zur Anwendung, wenn ein
dynamischer Reizverlauf auf den Rezeptor einwirkt. Moduliert nach
der jeweiligen variablen Dynamik vm(n...), welche als
STQ(v)-Parameter von der Membran gemessen wird, pflanzen sich die
AP's entlang des Axons mit STQ(v)-proportionaler Geschwindigkeit
vap(n..) fort. In gleicher Weise, wie in der Beschreibung zu Fig. 3c
die adaptive Anpassung der Zeitzählfrequenzen zu den Übergängen
2.1, 3.1, 4.1... dargelegt wurde, so ändern sich hier die
AP-Ioneneinstrom-Geschwindigkeiten v(t....) an der Synapse, die
Vesikel-Auffüllzeiten T(t...), die Quantität der Transmittermoleküle
in Vesikeln, ihre Bewegung in Richtung Vesikel-Gitter, die Struktur
dieses Gitters und viele andere Parameter der präsynaptischen und
subsynaptischen Strukturen.
Eine Synapse besitzt
Eigenschaften, die es ermöglichen, die Dynamik der einströmenden
AP's in vap-proportionale molekulare Zustandsveränderungen
umzusetzen. Diese wirken so, wie variable VTCP-Zeitzählimpulse in
Fig. 3c. Der Prozess kann verglichen werden mit variablem
Wasserdruck, der eine Turbine antreibt, durch welche ein Generator
variable Frequenzen erzeugt, die vom Druck und vom Tempo des
Wassers abhängen: erhöhter Wasserdruck steht für höhere Reizdynamik
am Rezeptor, für höhere axonale AP-Fortleitungsgeschwindigkeit vap,
und für höhere VTCP-Zeitzählfrequenz ƒscan in der Synapse (die sich
nicht nur auf das Tempo v(t) auswirkt, mit dem Vesikel sich
auffüllen, sondern auch auf viele andere synaptische Parameter).
Entsprechend dieser Prozesse werden die STQ(d)-Sequenzen
Td(1,2,3...) usw. in der Synapse mit vm-modulierten
Zeittaktfrequenzen ƒscan (1,2,3...) registriert; und als Konsequenz
daraus wird von dieser Zeitreihe die physikalische Struktur der
Synapse geprägt. Fig. 3d zeigt eine aktuell erfasste Zeitdaten-Reihe
32 30 22 23 20, die äquivalent ist zum registrierten Zeitmuster
Td(1,2,3,...), welche in der Synapse 24 eine spezifische moleku-
lar/biologische Spur hinterlässt. Die gezeigte früher erfasste
Zeitdatenreihe 30 29 22 24 19 in Fig. 3e entspricht jener
synaptischen Struktur, die durch oftmalige Reproduzierung bestimmter
Stimulus-Ereignisse und Zeitmuster Td'(1,2,3...) "geprägt" worden
ist.
Die manifestierte synaptische Td'- Struktur kann auch
als Fußstapfen-Sequenz betrachtet werden, die durch fortgesetzte
Lern-Prozesse und Wahrnehmungs-Erfahrungen erzeugt wurde, und die
z.B. als Referenz-Muster für das Ereignis "Nadelstich" dient. Wenn
eine neu erworbene Td-Fußstapfensequen - die durch gegenwärtige
Charakteristiken der Vesikel- Ansammlung und andere signifikante
zeitabhängige Parameter bestimmt wird - ungefähr mit dieser
bestehenden T'-Fußstapfenfolge (oder mit einem Teil davon!) Schritt
hält, dann wird in der synaptischen Struktur "Kovarianz" erkannt.
Dies resultiert in der Öffnung einer Vesikel- Andockstelle am
synaptischen Gitter und in der Freisetzung aller in einem Vesikel
enthaltenen Transmitter-Moleküle, worauf ein EPSP an der
sub-synaptischen Membran 25 produziert wird. Das Potential eines
EPSP entspricht jenen in Fig. 3f gezeigten
Wahrscheinlichkeitsdichte-Parametern, die für die momentan
bewertete Kovarianz signifikant sind. Wenn solche
Wahrscheinlichkeitsdichte-Parameter sich innerhalb eines bestimmten
Zeitintervalls zu einem bestimmten Schwellwert-Potential 27
addieren, wird ein AP 26 produziert. Dieses AP dient als
Bestätigung für das Ereignis " Nadel erkannt", und ruft einen
Muskel-Reflex hervor.
Der Vergleich aktueller
Verstreichzeitmuster mit früher erfassten Verstreichzeitmustern, wie
er in Fig. 3g gezeigt wird, findet in der Synapse fortgesetzt
statt. Jede erkannte Kovarianz einer neuen Zeitsequenz, die
durch besagte "zeitliche Auto-Adaption" registriert wird, setzt
eine Art von "Andock-Mechanismus" in Gang. Er initiiert jenen
Prozess, der vom Patentanmelder als "motorische Auto-Adaption"
bezeichnet wird, und welcher als der eigentliche "Motor" in
biologisch-chemischen Organismen bzw. Lebensformen verstanden werden
kann. Strukturen zeitlicher und motorischer Autoadaption, die auf
STQ-Quantisierung beruhen, existieren auch auf unterster
molekularer Ebene. Ohne Verstreichzeit-gestützte molekulare
Regelkreise wären koordinierte Zustandsveränderungen in
biologischen Systemen unmöglich. Dies trifft insbesondere auf
Bewegungen von Proteinen, auf die Erkennung und Kopierung des
genetischen Codes, und auf andere grundlegende Lebensprozesse zu.
Der Schaffung höherer biologisch/chemischer Ordnungen und komplexer
Systeme wie Synapsen oder Neurone geht die Existenz einer
STQ-quantisierenden molekularen Substruktur voraus, aus der einfache
Erkennungs- und Selbstorganisations-Prozesse von niedrigerem Niveau
abgeleitet werden. Tatsächlich gibt es unzählige Hierarchien von
auto-adaptiven Phänomenen auf verschiedenen Ebenen. Einfache
Phänomene auf molekularer Basis sind z.B. auch: das "Andocken" an
Rezeptormolekülen, die Bildung von Poren, Ionenkanälen und
subaxonalen Transportstrukturen (Mikrotubuli), und die Formierung
neuer Synapsen und axonaler Verzweigungen.
Demgegenüber
stellt die in der Beschreibung zu Fig. 4a - c gezeigte Erkennung von
Reiz- Signalverläufen durch synaptischen Zeitmuster-Vergleich (als
vegetativer Reflex oder als bewusste Wahrnehmung) ein Epiphänomen
bezüglich dessen dar. Jedes derartige auto-adaptive STQ-Epiphänomen
wird seinerseits von höherrangigen STQ- Epiphänomenen überlagert;
ein Beispiel dafür ist die Analyse komplexer
"Zeitmuster-Landschaften" auf Isomorphität. STQ- Epipänomene wie
Regelung des Wachstums, des Kreislaufes, der Körpertemperatur, der
Atmung, des Stoffwechsels, des Sehens, des Hörens, des Tastens, des
Riechens, der Bewegungskoordination usw. werden ihrerseits wieder
von noch komplexeren STQ-Szenarien übertroffen, wozu unter anderem
das Bewusstsein, das Denken, der freie Wille, das bewusste Handeln
oder auch die bewusste Zeitempfindung des Organismus zählen. In
allen diesen Fällen forscht das zentrale Nervensystem nach
konvergenten Zeitmustern, die wie einzelne Puzzle-Stücke zu einem
integrierten sensorischen Gesamtszenario zusammengesetzt werden.
Findet sich in irgendeiner Hierarchie innerhalb einer gewissen
"Latenzzeit" trotz intensiver "Suche" für eine Td-Sequenz kein
konvergentes Sub-Zeitmuster, so zeigt der Organismus chaotisches
Verhalten. Dieses Verhalten beschränkt sich auf jenen Teilbereich,
in dem der Kollaps aufgetreten ist. Sobald ein kovariantes
Zeitmuster gefunden ist, kehrt der geordnete Prozess der zeitlichen
und motorischen Auto-Adaption (und Auto-Emolution) zurück. Dies ist
auf triviale Weise mit einer Servolenkung vergleichbar, die
kurzfristig ausfällt. Das chaotische Verhalten wird jedoch
ebenfalls in seinem Ablauf als STQ-Zeitmuster quantisiert und in den
involvierten Synapsen auf eine Weise registriert, dass es trotz
einströmender AP's zu keiner Freisetzung von
Neurotransmitter-Substanzen kommt. Über subaxonale
Transportstrukturen (sog. Mikrotubili) strömen solche Informationen
als Transmittermoleküle in inverser Richtung entlang des Axons
zurück. Mikrotubili dienen zur Schaffung neuer Synapsen, und zur
synaptischer Anbindungen an solche Neuronen und neuronalen Netze,
in denen ebenfalls ein Kollaps eines Autoadaptionsprozesses
aufgetreten ist. Der Produzierung neuer Synapsen geht die
Entstehung von Dendriten voraus; das sind axonale Fortsätze, die aus
Neuronen hervorsprießen. Auf diese Weise regeneriert sich der
auto-adaptive neuronale "Feedback"- Mechanismus von selbst, und
jenes STQ-Zeitmuster, das während des kurzfristigen "chaotischen
Verhaltens" erfasst wurde, wird zu einer neuen Referenzbasis für
künftige Ereignisse. Auf diese Weise lernt der zentrale
Nervensystem (ZNS), neue Ereignisse und Erfahrungen zu registrieren;
und neue Zeitmuster einzubeziehen, die vorher unbekannt
waren.
Fig. 5 zeigt eine Konfiguration, in der die beschriebene erfundene Methode angewandt wird, um einen autonomen selbst-organisierenden Mechanismus (insbes. einen Roboter) zu generieren, bei dem die STQ-Quanten mittels mechanistischer Sensor-Technologie und Elektronik-Schaltkreise erworben werden. Im Unterschied zu Fig. 4a - f werden im gezeigten Fall beinahe ausschließlich STQ(i)-Verstreichzeiten zusammen mit STQ(v)- Verstreichzeiten (die für die Messung der relativen Momentangeschwindigkeit benötigt werden) quantisiert. Die mit Tw(1,2,3....n) bezeichneten Zeitdatenströme resultieren aus den vm-adaptiven STQ(i)-Verstreichzeitmessungen. Nichtsdestoweniger wäre es aber vorteilhaft, auch STQ(d)- Quanten zu erfassen, die zur Verifizierung von erworbenen Tw-Zeitdatenströmen dienen können. Im Gegensatz zu molekular/biologische Organismen ist es in mechanistischen Syste- men kaum möglich, eine verhältnismäßig große Anzahl von Sensoren auf engstem Raum neben- einander zu placieren. Daher ist es erforderlich, aus der verfügbaren mechanistischen Sensor- Technologie soviel STQ- Verstreichzeiten als möglich zu gewinnen, um eine ausreichende große Referenzbasis für die nachfolgende statistische Analyse zu erlangen. Wie bereits in der Beschreibung zu Fig. 3a ausgeführt wurde, fallen bei multiplen STQ(i)-Quantisierungen Zeitdaten parallel und gleichzeitig an, weshalb diese Daten auch in paralleler Weise verarbeitet werden müssen.
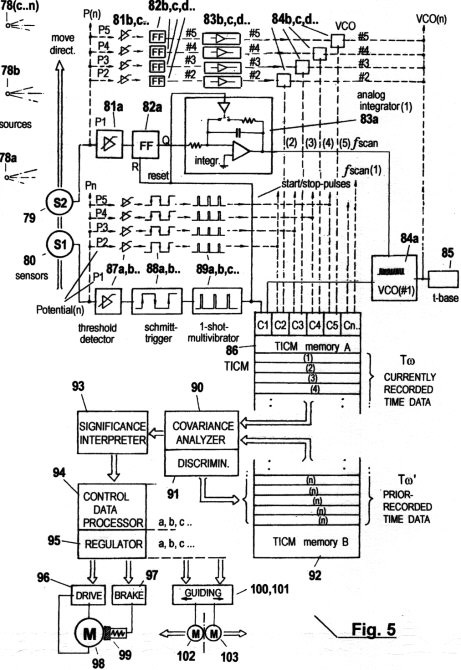
Die
Konfiguration zeigt ein Blockschaltbild eines mobilen autonomen
Roboters mit der angeeigneten Fähigkeit, Bewegungsphasen in
auto-adaptiver Weise zu reproduzieren und das Timing seiner
Eigenbewegung durch fortgesetztes Abtasten und Erkennen seiner
physikalischen Umgebung zu optimieren. Das Robotiksystem ist mit
gleichartigen benach- barten Sensoren 79 und 80
bestückt, welche analoge Ausgangssignale liefern, die mit
Schwellwertdetektor-IC's 81a, b, c, d ,e.. und
87a, b, c, d, e... verbunden sind. Bewegt sich Sensor
79 (der "V-Sensor") in der bezeichneten Richtung entlang der
korrespondieren- den äußeren Signalquelle 78a, so durchbricht dessen
Signalamplitude zunächst das niedrigste Potential P1, das vom
Schwellwertdetektor 81a bestimmt wird (s. Beschreibung zu Fig. 2b).
Dadurch wird das Flip-Flop 82a getriggert (Output = H). Ein
Schmitt-Trigger-IC sowie ein Monoflop-IC sollte vorgeschaltet sein,
um bei jedem Phasen- durchgang kurze Impulse abzugeben. Der
nachgeschaltete rücksetzbare Präzisions- Integrator(1) 83a
liefert nun ein stetig ansteigendes analoges Output-Signal, das die
Frequenz des programmierbaren Oszillators (VCO) 84a
moduliert. Diese Frequenz ist mit dem Input eines digitalen TICM
(Vielfach-Zeitzähl- und Speicher-IC) 86(C1) verbunden und
bildet die aktuelle vm-adaptive Zeitzählfrequenz ƒscan(1) (s. auch
Fig. 3b, 3c). Der Integrator (1) 83a dient demnach zur
STQ(v)- Quantisierung. Er misst die Verstreich- zeit Tv(1) in Form
eines Potentialanstiegs, der über den VCO(#1) 84a in besagte
Zähl- frequenz ƒscan(1) umgewandelt wird, welche anti-proportional
zu den relativen Geschwindig- keiten vm(n..) ist, mit der sich das
Robotiksystem gegenüber der räumlichen physikalischen Umgebung
bewegt.
Gelangt der benachbarte Sensor 80 (der
"W-Sensor") in den Wahrnehmungsbereich der Signalquelle 78a,
so durchbricht seine Signalamplitude zunächst das vom
Schwellwert-detektor-IC 87a bestimmte niedrigste
Potential P1 (s. Beschreibung zu Fig. 2b). Als Ergebnis produziert
die steigende Flanke des anschließenden Schmitt-Trigger-IC
88a einen Impuls im nachgeschalteten Monoflop-IC 89a,
der im TICM 86 (C1) die STQ(i)-Quantisierung der vm-
adaptierten Verstreichzeit Tw(1) startet. Da gleichzeitig ein
Reset-Impuls an das Flip-Flop 82a geht und daher der
analoge Pegel am Output des Integrators(1) 83a
gehalten wird, verbleibt die Zählfrequenz ƒ(1) als momentane
vm-abhängige Zeitzählbasis ƒscan (1) am Output des TICM
86(C1) solange unverändert, bis der nächste STQ(v)-Parameter
quantisiert wird. Diese Quantisierung findet statt, wenn die
Signalamplitude des Sensors 79 unter das vom
Schwellwertdetektor-IC 81a festgelegte Potential P1 fällt
(wobei das Flip-Flop 82a von der fallenden
Signalflanke getriggert wird), oder wenn der Sensor 79 in den
Wahrnehmungs- bereich einer weiteren Signalquelle 78b,c,d,e..
gelangt.
Gleichzeitig wird dabei über IC's 87a, 88a
und 89a erneut ein Impuls produziert, der die Messung der
Verstreichzeit Tw(1) im TICM 86(C1) stoppt und die gezählten
vm- modulierten Zeitimpulse im Zeitdatenspeicher (C1) ablegt. Im
Memory-Bereich C1 sind jene Tw-Verstreichzeiten gespeichert, die
sich auf das niedrigste Potential P1 beziehen, beispiels- weise
Tw(1), Tw(8), Tw(15) usw. Die Quantisierung aller auf die höheren
Potentiale P2, P3, P4, P5 usw. bezogenen STQ-Verstreichzeiten wird
in derselben Weise abgewickelt wie für P1. Passiert die
Signalamplitude aus Sensor 79 die von den Detektor-IC's
81b, c, d, e... bestimmten Schwellwert-Potentiale P2, P3,
P4, P5.., so triggern nacheinander die Outputs der Flip-Flop's
82b, c, d, e.. auf = H, und die nachgeschalteten
Integratoren 83b, c, d, e... liefern stetig
ansteigende analoge Output-Pegel, welche ihrerseits die Frequenzen
ƒscan (von den VCO's 84b, c, d, e.. produziert) solange
stetig vermindern, bis die Signalamplituden aus Sensor 80
die von den Detektor-IC's 87b, c, d, e.. bestimmten höheren
Schwellwert- Potentiale P2, P3, P4, P5.. erreichen, sobald dieser
Sensor in den Wahrnehmungsbereich der Signalquelle 78a
gelangt.
Daraufhin werden die Schmitt-Trigger 88b, c, d,
e.. aktiviert, und die Monoflops 89b, c, d, e...
produzieren Impulse, die im TICM 86(C2,C2,C3,C4 ...Cn) die
Erfassung der vm-adaptierten Verstreichzeit-Daten Tw(1,2,3,4....n)
starten. Die Registrierung dieser Daten geschieht während die
momentanen vm-adaptierten Zeitzählfrequenzen ƒscan (1,2,3,4,..n)
gelten, weil die gleichzeitig an die Flip-Flops 82b, c, d, e..
ergangenen Reset-Impulse die Output-Pegel der Integrator-IC's
83b, c, d, e.. halten, womit die entsprechenden Output-
Frequenzen ƒ(1,2,3,4...n) an den VCO's 84b, c, d,
e...festgelegt werden. In derselben Weise erfolgen die
fortgesetzten Quantisierungen weiterer Tw-Verstreichzeiten, wenn
sich die Sensoren 79, 80 entlang weiterer
Signalquellen 78b, c, d, e... bewegen. Im TICM 86
(C1...n) werden alle quantisierten STQ(i)-Zeitdaten abgelegt. Im
Speicherbereich C2 (s. die korrespondierende Fig. 2b) befinden sich
die auf den Wahrnehmungsbereich
(Potential) P2 bezogenen Verstreichzeiten Tw(2), Tw(7), Tw(14)...,
im Speicherbereich C3 die auf die höhere Potentialstufe P3
bezogenen Verstreichzeiten Tw(3), Tw(6), Tw(13)..., im
Speicherbereich C4 die auf die nächsthöhere Potentialstufe P4
bezogenen Verstreichzeiten Tw(4), Tw(5), Tw(12)..usw. Die im TICM
86 einströmenden aktuellen Tw-Sequenzen werden demnach durch
die aktuelle Bewegung des sensor-gekoppelten autonomen Mechanismus
(z.B. "Roboter-Vehikel") entlang einer bestimmten Bahn generiert.
Im gezeigten Beispiel weichen die Positionen der Sensoren bezüglich
der Positionen der externen Signalquellen (den physikalischen
Umgebungen) zeitlich ab.
Im Falle absoluter physikalischer
Invarianz zwischen dem autonomen Vehikel und den Umgebungen (auch
ident mit "absoluter Synchronität") kann weder ein STQ-Parameter
noch eine Tw-Sequenz erworben werden. Tritt keine solche
physikalische Invarianz auf, dann ist es für das autonome Vehikel
möglich, seine eigene Bewegung entlang der Bahn durch den stetigen
Vergleich aktuell erfasster STQ-Zeitdatenmuster Tw(1,2,3..n) mit
früher erfassten STQ-Zeitdatenmuster Tw'(nnnn) zu erkennen; und es
ist auch möglich die erkannten Bewegungen fortgesetzt in
auto-adaptiver Weise zu optimieren. Voraussetzung dafür ist, dass
das Vehikel über ein entsprechendes Antriebs- und Bremssystem
verfügt, das durch Daten kontrolliert wird, die mittels
fortgesetzten statistischen Zeitdaten-Analysen errechnet werden.
Man vergleiche dazu auch Fig. 3d und 3e: Sobald die
Regressionskurve
einer aktuellen, im TICM 86 gespeicherten
Zeitdaten-Sequenz Tw (1,2,3...), mit der
Regressionskurve
einer zeitlich zurückliegenden, durch eine frühere gleichartige
Bewegung entlang derselben erfassten Zeitdaten-Sequenz Tw'(nnn)
konvergiert, wird das Antriebssystem 98 (sowie das
Bremssystem 99) durch Impulse 96, 97 aktiviert, die
das Vehikel dazu veranlassen, seine Bewegungsverläufe entlang den
externen Signalquellen 78a, b, c, d, e... in einer Weise zu
vollziehen, daß der aktuelle Bewegungsverlauf mit jenem früheren
Bewegungsverlauf annähernd räumlich und zeitlich isomorph ist, aus
dem die Zeitdaten-Sequenz Tw'(nnnn) hergeleitet wurde. Zu diesem
Zweck sind das TICM 86, in das die aktuellen
Zeitdaten einfließen, sowie die Speichereinheit
92, in der die früher erfassten Zeitdaten Tw'(nn)
gespeichert sind, mit einem Kovarianz- Analysator 90 und
einer Diskriminator- Logik 91 verbunden, welche die einzelne
Zeitdaten verifiziert und auf Plausibilität prüft. Ungültige
Zeitdaten werden gelöscht oder so interpoliert, dass kein Ausfall
eines datengestützten Regelkreises auftreten kann. Analysator
90 und Diskriminator 91 suchen den Speicher 92
mit sehr hoher Frequenz fortgesetzt nach annähernd übereinstimmenden
Zeitdatenmustern ab. Signifikante Datensequenzen werden an den
Interpreter 93 weitergeleitet, welcher über die
jeweilige Wahrscheinlichkeitsdichte und das Ausmaß der Kovarianz
entscheidet. Besteht ausreichende Übereinstimmung, so errechnet der
Prozessor 94 die entsprechenden Aktivierungsparameter
Tempoverteilung zur Erzielung eines isomorphen Bewegungsverlaufes.
Diese Daten gelangen an das Steuerungsmodul 95, wo sie in
Impulse 96, 97 für das Antriebs- Bremssystem und 98,
99 umgesetzt werden.
Es ist ferner von Vorteil, in diesen
Prozess auch energetische Impulse für ein Lenk- und
Gegenlenksystem
100, 101, 102, 103 einzubeziehen, das auf denselben
beschriebenen Funktionsprinzipien aufbaut und das für die
Einhaltung des ebenfalls von besagten Tw'- Zeitmustern festgelegten
räumlichen Bewegungsverlaufes sorgt. Voraussetzung für eine
einwandfreie Funktion einer solchen Anordnung ist die Verwendung
extrem schneller Prozes- soren zur Bedienung der Teilsysteme 90,
91, 93, 94 und 95. Der aktuelle Bewegungsverlauf des
autonomen Vehikels kann nur dann annähernd isomorph zum referenten
Bewegungs- verlauf werden, wenn die Erkennung der signifikanten
Tw'(nnnn)-Sequenzen die Registrierung und Analyse der aktuellen
Tw(1,2,3...)- Sequenzen (Ist-Werte), die Errechnung der
Regelparameter und die Applizierung der Energie-Impulse
96, 97 in Echtzeit erfolgen. In diesem Fall zeigt das
Vehikel eine Verhaltensweise ähnlich wie ein "Servo-Regelkreis"
bekannter Art. Diese Affinität wird auch durch die Tatsache
bestätigt, dass durch einfaches Erhöhen oder Verringern der
Basisfrequenz fn der Zeittaktquelle 85 der gesamte zeitliche
Verlauf aller einzelnen Bewegungsphasen völlig synchron
beschleunigt oder verlangsamt werden kann. Jedem äußeren Eingriff,
der den Bewegungsverlauf zu ändern oder zu stören versucht, wird
automatisch vom Antriebsmechanismus des autonomen Vehikels
entgegengewirkt. Deshalb ist ein autonomer Mechanismus, der nach
diesen Prinzipien arbeitet, mit einem "lebenden Organismus"
vergleichbar.Da in den System-Elementen 90, 91, 93, 94 und
95 das Bestreben programmiert ist, die Analyse und
Interpretation der erfaßten Zeitparameter fortgesetzt zu verbessern
(z.B. nur solche Tw'(nnnn)- Zeitdaten zu erlauben, die der kürzesten
und effizientesten Linienfolge zuzuordnen sind), existiert in
einem solchen Mechanismus nicht nur ein Bestreben nach zeitlicher
und motorischer Auto-Adaption, sondern auch eine "Optimierungs-
Tendenz". (Dies ist auch in molekular/biologischen Strukturen von
Organismen inhärent (s. auch Beschreibung zu Fig. 4a -
4f. Das System ist auch imstande, Prioritäten zu setzen, sowie sich für
Tw'-Zeitdatensequenzen zu entscheiden, die mit einer anderen
Regressionskurve korrespondieren, falls z.B. eine irreguläre
Bahnabweichung erkannt wird, die vom Steuerungsmodul 95
nicht mehr stabilisiert werden kann; daraufhin strebt das Vehikel
einem neuen Bewegungsverlauf und einer neuen Tempoverteilungskurve
(Timing) nach. Im Memory des TICM 86 können beliebige
alternative Bewegungs-Szenarien in Form von Tw'- Zeitdatenmustern
gespeichert sein, auf die "zugegriffen" werden kann, wenn es eine
bestimmte Kursabweichung erfordert. Auf diese Weise sind
Crash-Situationen bereits im Ansatzpunkt zu erkennen und können
verhindert werden, indem das Vehikel in der Lage ist, in autonomer
Weise entgegenzuwirken.
Das System gerät nur dann außer
Kontrolle ("chaotischer Zustand"), wenn keinerlei aus früher
registrierten Tw-Sequenzen abgeleitetes Regressionskurven-Segment
gefunden worden ist, welches mit irgendeinem
Regressionskurven-Segment konvergent ist, das aus aktuell
registrierten Tw-Sequenzen abgeleitet ist. Dieser Prozess wird vom
Anmelder als "motorische Auto-Adaption" oder als "Auto-Emulation"
bezeichnet. Um zeitlich/räumliche Zustands- veränderungen der
physikalischen Umgebung aus subjektiver Sicht des autonomen Systems
zu identifizieren, genügt es in den meisten Fällen nicht, externe
räumliche Strukturen, Landmarken und Lichtverhältnisse mittels
optischer oder fotoelektrischer Sensoren
passiv abzutasten. Üblicherweise ist es auch nötig, die
Höhenabweichungen mittels
Neigungssensoren, Wegunebenheiten mittels Druckaufnehmer oder
Beschleunigungs-Sensoren, stationäre akustische Quellen mittels
Mikrofone, Gradienten mittels Magnetfeldsensoren;
sowie Positionen mittels GPS-Empfänger abzutasten, um genügend
STQ-Zeitparameter als Referenzbasis zu gewinnen.
Alle diese
aufgenommenen Tw(n)-Zeitdatenströme werden im Memory des TICM
gespeichert. Daraus kann man folgern, dass die Lern- und
Selbstorganisations-Fähigkeit
eines Organismus (bzw. eines autonomen auto-adaptionsfähigen
Mechanismus) proportional zur Menge aller verfügbaren Sensoren bzw.
zur Anzahl aller dem Autoadaptionsprozess verfügbar
gemachten STQ- Parameter zunimmt. Eine weitere wichtige Erkenntnis
ist, dass es kein Timing ohne begleitende Zeitnahme (=
STQ-Quantisierung) gibt. Auto-adaptive Prozesse und Mechanismen der
beschriebenen Art werden für viele zukünftige Aufgaben im Bereich
der Hochtechnologie, z.B. für die Entwicklung autonomer
Robotersysteme, unverzichtbar sein. Ein Beispiel dafür ist
folgendes: Ein Automobil, das sich im Straßenverkehr in autonomer
Weise sicher und effizient fortbewegen soll, muss imstande sein,
seitliche und frontale Abstände sowie fixe Geschwindigkeitsphasen
einzuhalten. Mehr noch: es muss auch fähig sein, selbsttätige
Überholmanöver auszuführen, und Gefahrensituationen antizipierend
zu erkennen und zu vermeiden. Dies ist nur möglich, wenn der
On-Board-Computer im Fahrzeug mit einer Vielzahl verschiedener
Sensoren zur Erfassung vielfältiger Signalquellen gekoppelt ist, und
wenn er über eine sehr schnelle und effiziente Hardware und Software
verfügt, welche jene STQ-Zeitdaten, die für die Autoadaption
benötigt werden, annähernd in Echtzeit verarbeiten kann. Zukünftige
Typen von Mikroprozessoren könnten um eine hardwaremäßige Struktur
erweitert werden, welche die beschriebenen Funktionen
vollbringen.
Fig. 6a zeigt die Konfiguration einer
einfachen Verkörperung eines Aspekts der Erfindung, bei der die in
Fig. 2a - 2c erläuterten STQ(v)-, STQ(i)- und
STQ(d)-Quantisierungsmethoden zur Erkennung von räumlichen Profilen
oder Strukturen angewandt werden. Im hier gezeigten Anwendungsfall
soll ein Roboterarm, auf zwei benachbarte Metallsensoren 104,
105 im Abstand B installiert sind, fähig sein, das Profil des
Metallstabes 106 von anderen zu unterscheiden, während er
sich mit beliebiger Geschwindigkeit an beliebigen Stäben 106,
107, 108 usw. entlang bewegt.
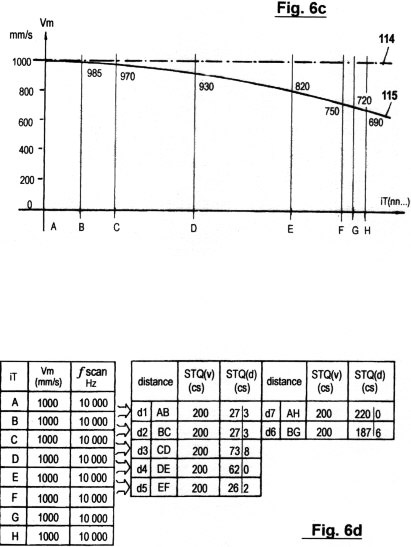
Bewegt sich der Sensorkopf in der Höhe h entlang der bezeichneten Richtung, so gelangen der v- Sensor 104 (S2) und dann der w-Sensor 105 (S1) nacheinander in jenen niedrigen sensitiven Bereich, der hier als "Wahrnehmungsintensitätszone 3" bezeichnet ist. Der niedrigste Schwellwert P1 wird von der Signal-Amplitude durchbrochen, und die im Wesent- lichen aus den Elementen 81, 82, 83, 84, 85, 86, 87, 88, und 89 bestehende Aquisitions-Logic 109 (s. auch Fig. 5) beginnt mit dem Erwerb vm- modulierter STQ(i)- STQ(d)- Zeitsequenzen Tw(1,2,3...n) und Td(1,2,3...n), die im TICM-Memory (A) 110 gespeichert werden. Derselbe Zeitdatenerfassungs-Prozess wiederholt sich, wenn die Sensoren 104, 105 auf die nächsthöheren Wahrnehmungsbereichszonen 2 und 3 treffen und die Signalamplituden die in den Schwellwertdetektoren eingestellten Potentialwerte P2 und P3 durchbrechen. Um jene Metallstäbe 106, die das charakteristische Profil aufweisen, im Analysator 112 eindeutig zu identifizieren, müssen die Tw,Td-Zeitdatenströme, welche ins Memory 110 einfließen, fortgesetzt mit jenem signifikanten Tw',Td'-Zeitdatenmuster (B) 111 verglichen werden, das als "Referenzmuster" vorprogrammiert worden ist. Ungültige oder irreguläre - in der Diskriminator-Einheit 113 erkannte - Zeitdaten werden sodann gelöscht oder korrigiert. Diese Einheit ist auf die Fähigkeit programmiert, die Zuordnung und Verarbeitung der Daten automatisch zu verbessern (d.h. die Zeitdaten in auto-adaptiver Weise zu überprüfen und zu verifizieren) wie dies schon in bezug auf Fig. 5 beschrieben wurde. Wurde ein Profil als solches erkannt, so gibt der Analysator 112 ein Bestätigungssignal an eine Aktuator-Einheit des Roboters weiter, die dann einen Mechanismus in Gang bringt, der die identifizierte Metallschiene vom Boden abhebt, auf ein Laufband aufsetzt usw.
Fig. 6b - e zeigen verschiedene Diagramme und Tabellen bezüglich Fig. 6a
Fig. 6b zeigt ein Sensorfeld-Diagramm des abgetasteten Metallprofils 106. Die Messung seiner Dimensionen d1....d7 resultiert ausschließlich aus STQ- Quanten, d.h. in der Zeit- domäne. Identifikation der Profile sind 3 Empfindlichkeitszonen (P1, P2 und P3) vorgesehen, die in Schwellwert-Detektoren gespeichert werden. Zu den Phasendurchgängen (iT)A, (iT)B, (iT)C, (iT)D, (iT)E, (iT)F, (iT)G und (iT)H werden digitale Präzisionszeitmesser gestartet oder gestoppt. Da die variable Zeitzählfrequenz ƒscan, mit der diese Zeitmesser zählen, automatisch an die Abtast-Geschwindigkeit vm angepasst (bzw. moduliert) wird (siehe auch Fig. 3a - g, und Fig. 5), korrelieren die tatsächlichen Dimensionen d1....d7 weitgehend mit den Tw,Td- Verstreichzeiten, die bereits im Memory 110 gespeichert sind. Wie aus der Zeichnung ersichtlich, erhält man die Distanzen AB-(d1) und BC-(d2) aus STQ(d)-Verstreichzeiten, die Distanzen CD-(d3), DE-(d4), EF-(d5) sowie BG-(d6) und AH-(d7) aus STQ(i)- Verstreichzeiten. Einmal mehr wird vom Patentanmelder bekräftigt, dass alle (iT)n... flüchtige Phasendurchgänge darstellen, und niemals "Zeitpunkte" nach klassischem physikalischem Verständnis.
Fig. 6c zeigt die vm-Diagramme zweier Bewegungsverläufe der Sensoren S1 und S2 entlang des abgetasteten Metallprofils. Im einen Fall bewegt sich der Roboterarm, auf dem die beiden Sensoren montiert sind, mit einer konstanten Geschwindigkeit von 1000mm/s über dem Profil (strichpunktierte Linie 114). Im anderen Fall wird die Geschwindigkeit des Arms von 1000mm/s beim ersten Phasendurchgang A bis auf 690mm/s beim letzten Phasendurchgang H abgesenkt. Diese Absenkung ist nicht linear, sondern folgt dem Verlauf der Linie 115.
Fig. 6d zeigt eine fiktive Frequenz- und Zeitdatentabelle zu Fig. 6c bei einer gleichbleibenden relativen Geschwindigkeit von vm=1000m/s an allen Phasendurchgängen (iT) A...H. Daraus ergibt sich eine vm-modulierte Zeitzählfrequenz ƒscan von 10 kHz während des gesamten Abtastvorganges. Da im vorliegenden Fall die STQ(v)-Verstreichzeitmessung mit einem fixen (primären) Quellen-Zeittakt von 200cs/b erfolgt, resultieren aus dem Scanning- Prozess vm-adaptierte STQ(d)-Verstreichzeiten von 273cs, 273cs, 738cs, 620cs und 262cs für die Distanzen AB, BC, CD, DE und EF sowie vm-adaptierte STQ(i)-Verstreichzeiten von 1876cs und 2200cs für die Distanzen BG und AH. Diese aus STQ(d)- und STQ(i)-Verstreichzeiten bestehende aktuelle Tw,Td-Sequenz wird im Analysator 112 mit der referenten gespeicherten Tw',Td'-Sequenz 270cs, 270cs, 740cs, 620cs, 260cs, 1880cs, 2200cs verglichen, welche als signifikantes Zeitmuster für dieses Metallprofil dient, das bereits im Memory 111 gespeichert ist. Entscheidet der Analysator, daß "Übereinstimmung" besteht, so wird ein Bestätigungssignal an eine Aktuator-Einheit ausgegeben. Der Analysator verfügt über Komparatoren und/oder "Fuzzy-Logic"-IC's, welche die Streuungen in Grenzbereichen ignorieren (z. B. Dezimalstellen aufrunden). Abgesehen von diesen Korrekturmaßnahmen lassen sich auch Toleranzen, Plausibilitätskriterien und Zuordungskriterien softwaremäßig programmieren.
Fig. 6e zeigt dieselbe Frequenz- und Zeitdatentabelle wie Fig. 6d, jedoch mit variablem Verlauf der Abtast- Geschwindigkeit(vm). Die Relativgeschwindigkeit von 1000mm/s an Phasendurchgang (iT)A nimmt auf 690 mm/s beim letzten Phasendurchgang (iT)H ab. Die vm- Absenkung ist nicht linear. Entsprechend des Verlaufes des Graph 115 werden zu den Phasendurchgängen (iT)A, B, C, D, E, F, G, H die einzelnen Momentangeschwindigkeiten (vm1,2,3..) 1000; 985; 970; 930; 820; 750; 720 und 690mm/s gemessen. Die besagte vm- adaptive Modulation der Zeitzählfrequenz ƒscan produziert zu den Phasendurchgängen die Frequenzwerte 10; 9,85; 9,70; 9,30; 8,20; 7,50; 7,20 und 6,90kHz, mit denen die STQ(i)- und STQ(d)-Verstreichzeiten quantisiert werden. Da die STQ(v)-Quantisierungen ebenfalls mit einem fixen Quellen-Zeittakt von 200cs/b erfolgt, resultieren für die Distanzen AB, BC, CD, DE, EF, BG und AH dieselben Tw,Td-Verstreichzeit-Sequenzen, wie in Tabelle Fig. 6d. Daraus ist ersichtlich, da die Erkennung des besagten Metallprofils sowohl bei linearer als auch bei nichtlinearer vm-Tempoverteilung gewährleistet ist.
Fig. 7a - d zeigen verschiedene Sensor-Konfigurationen zur
Quantisierung von STQ(v)- Verstreichzeiten bzw. zur Erfassung der
Relativgeschwindigkeits-Parameter (vm). Die ersten drei Konfigurationen
zeigen Sensoranordnungen für 2-dimensionale Erfassung von externen
Ereignissen, Fig. 7d zeigt eine besondere Konfiguration anwendbar für
beliebige drei- dimensionale Erfassung der physikalischen
Umgebungen.
Fig. 7a zeigt eine Sensoranordnung, bei dem eine
Halterung, die S1 und S2 in Entfernung b auf gleicher Achse trägt, sich in
der gekennzeichneten Richtung entlang einer willkürlichen Bahn bewegt,
bzw. sich auf einer Oberfläche um ein räumliches Zentrum A in Äquidistanz
zu S1 (V-Sensor) und S2 (W-Sensor) dreht. Dieses Sensor-System verfügt
nur über einen Freiheitsgrad.
Fig. 7b zeigt eine
Sensoranordung, bei dem eine Trägerfläche, die auf gleicher Achse zwei
V-Sensoren S2 und einen in Äquidistanz b zu beiden befindlichen W-Sensor
S trägt, sich beliebig in einer der beiden gezeigten 180°
entgegengesetzten Richtungen entlang einer willkürlichen Bahn bewegt,
bzw. beliebig um einen im Raum befindlichen Punkt A dreht, der
äquidistant zu beiden V-Sensoren S2 ist. Die beiden in Fig. 7a und Fig.
7b gezeigten Sensoranordnungen sind für die meisten verkehrstechnischen
Robotik-Anwendungsgebiete ausreichend.
Fig. 7c zeigt eine
Konfiguration mit einer Anzahl gleichgearteter V-Sensoren S2, die als
Segmente rund um einen zentralen W-Sensor S1 auf einer kreisförmigen
Trägerfläche mit Radius b gruppiert sind. Bei dieser Konstellation kann
sich die Trägerfläche in irgendeiner Richtung auf willkürlicher Bahn
bewegen, oder sich beliebig um einen in irgendeinem Sensor- Abstand im
Raum befindlichen Punkt drehen. Diese Sensoren-Anordnung verfügt daher
über 2 Freiheitsgrade.
Fig. 7d zeigt eine
Sensor-Konfiguration mit einer Vielzahl von V-Sensoren S2, die als Seg-
mente auf einer kugelförmigen Trägerfläche im Radius b rund um einen
zentralen W-Sensor S1 gruppiert sind. Die Sensor-Anordnung kann sich zu
beliebiger Position im dreidimensionalen Raum bewegen oder in beliebiger
Richtung um ein festes räumliches Zentrum A mit beliebigem Abstand zu den
Sensoren rotieren. Diese Konfiguration verfügt über 3 Freiheitsgrade. Die
in Fig. 7c und Fig. 7d gezeigten Sensor-Anordnungen kommen in erster Linie
für autonome Erkundungsroboter oder Flugobjekte in Frage, bei denen
Antriebs-Impulse in beliebiger Richtung appliziert werden können (z.B.
mittels Steuerungsraketen).
-------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
Fig. 8a - f zeigen die
Konfiguration und Funktionsprinzipien einer weiteren Verkörperung der
gegenständlichen Erfindung, bei der die in Fig. 2a, b, c beschriebenen
STQ-Quantifizierungs- Methoden zur Schaffung eines autonomen lernfähigen
selbstorganisierenden Trainingsroboters für den Einsatz im Sport dienen;
einen sogenannten "elektronischen Hasen". Dieses System besitzt autonome
Antriebs-, Lenkungs- und Bremsmechanismen sowie einen Analysator, der
die aktuell registrierten vm-adaptiven STQ(i)- und STQ(d)-Zeitstrommuster
Tw,Td(1,2,3...) fortgesetzt mit früher registrierten vm-adaptiven STQ(i)-
und STQ(d)-Zeitstrommuster Tw',Td'(1,2,3...) vergleicht, welche als
Referenzmuster dienen. Es ist dadurch imstande, einen vom Benutzer
vorgegebenen Bewegungsvorgang zu reproduzieren und zu optimieren; die
idealen Routen und die Tempoverteilungen automatisch zu finden; Abstände
und Zeiten einzuhalten; Gefahrensituationen zu erkennen und davor zu
warnen; und die Eigenbewegung sowie Informationen über Geschwindigkeit,
Rundenzeiten, Teilzeiten, Gesamtzeiten usw. auf einem Monitor
darzustellen. Zusätzlich ist es imstande, diese Daten wahlweise optisch
oder akustisch auszugeben.
Fig. 8a zeigt einen derartigen
Trainingsroboter 116 vor einem Skilangläufer
117. Das dargestellte Roboter-Vehikel verfügt über ein
Kufen-Fahrgestell, das die Fortbewegung schneebedecktem Boden
ermöglicht. Um imstande zu sein, einem menschlichen Skifahrer
entlang einer Langlaufspur zu folgen, hat es entsprechend
manövrierfähig zu sein. Es muss auch fähig sein, eine Langlaufloipe
selbsttätig zu spuren, wenn die alte Spur vom Schnee zugeweht wurde
und daher nicht mehr sichtbar ist. Der Trainingsroboter ist speziell
auch als Hilfsmittel für blinde Skiläufer geeignet. Das autonome
Vehikel erkennt skiläuferische Kriterien für blinde Skilangläufer
und ruft Hinweise, Berichte und Warnungen mit Hilfe von
Sprachsynthese. Dies befreit den Anwender vom Gefühl, auf Hilfe
angewiesen zu sein, und macht ihm mehr Freude. Das Robotervehikel
116 verfügt über eine hohe Anzahl von Sensoren und
elektronischen Einheiten, wie sie in Fig. 5 beschrieben wurden. Es
vollbringt dieselben darin aufgezeigten Auto-Emolutions-,
Auto-Adaptions- und Auto-Optimierungs-Prozesse, wobei häufig
verschiedene Aufgaben gleichzeitig durchgeführt werden. Es bezieht
vm- adaptierte STQ(i)- und STQ(d)-Verstreichzeitmuster aus einer
Vielzahl von Sensoren, vergleicht diese Muster mit den
entsprechenden Referenzzeitmustern, wählt die signifikanten
Zeitdaten aus und analysiert und errechnet Parameter für die
diskreten Energie-Impulse zur Steuerung der Antriebs-, Brems- und
Lenkmechanismen.
Im folgenden Teil werden die
wesentlichsten Komponenten des Systems unter Einbeziehung von drei
spezifischen Typen von Sensoren (optische S., Magnetfeld- und
GPS-Positions-Sensoren) behandelt.
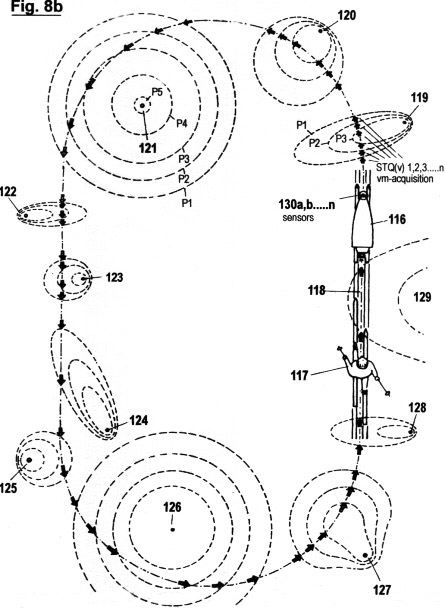
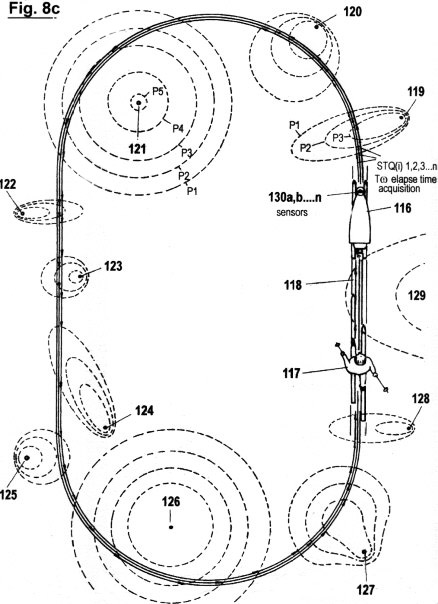
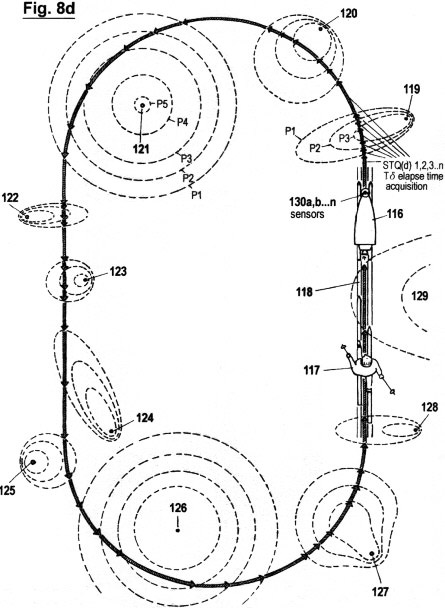
Fig. 8b - d zeigen die Registrierung von
STQ(v)-, STQ(i)- und STQ(d)-Verstreichzeiten (passend zu Fig. 8a)
bei Verwendung optischer oder akustischer Sensoren. Die fundamentalen Funktionsprinzipien sind zum größten Teil bereits in den
Beschreibungen zu Fig. 2a, b, c und Fig. 5 enthalten. In besagten
Zeichnungen bewegt sich der Trainingsroboter ("der elektronische
Hase") 116 mit variabler Geschwindigkeit vor einem
Skilangläufer 117 in der Loipe 118. Entlang
der Spur befinden sich in willkürlicher Anordnung optische oder
akustische Signalquellen 119, 120, 121, 122, 123, 124,
125, 126, 127, 128 und 129, die von ent-
sprechenden Sensoren 130a, b,...n wahrgenommen werden. Bei
jedem Phasendurchgang zu den Schwellwertbereichen P1, P2, P3, P4,
P5 usw. werden die besagten STQ(v)-, STQ(i)- und
STQ(d)-Verstreichzeiten erfasst. Sie ergeben das aktuelle
vm-adaptive Tw,Td(1,2...n)- Zeitdatenmuster, das im TICM gespeichert
wird. Für die prinzipielle Funktionsweise ist es nicht von
wesentlicher Bedeutung, ob die Signalquellen dort stationär
angeordnet sind (z.B. könnten sie Scheinwerfer sein, welche die
Piste für abendliche Events beleuchten). "Signalquellen" können
auch aus wechselnder Helligkeit bei Tageslicht oder aus Kontrast-
und Farbunterschieden nahe Bäumen, Masten, Gebäuden, Abhängen oder
signifikanten Landmarken resultieren. Scheinwerfer könnten sogar am
Schulungs-Roboter selbst installiert werden, wobei die
optosensorische Erfassung des reflektierten Lichts und die
Evaluierung der Licht-Strukturen der räumlichen Umgebungen zur
Erkennung der Eigenbewegung genutzt werden kann. Die gleiche
Anordung wäre auch mit Ultraschallgeber und -sensoren machbar.
Akustische Signalquellen können anderseits ebenso natürlichen
Ursprunges sein, wie etwa das Rauschen eines Baches neben der Loipe,
oder ein Wasserfall usw. Generell kann jedwede flüchtige
Kombination von Licht und Schatten oder irgendeine Geräuschquelle
signifikant für die Erkennung eines bestimmten Objekts sein. Die
Bestimmung der individuellen Identität eben dieses Objekts
geschieht durch Vergleich von vm-adaptiv registrierten STQ(i)- und
STQ(d)-Verstreichzeit-Mustern, die für jedes einzelne Objekt
stehen.
Um die Beschreibung und Veranschaulichung zu
vereinfachen, wird in Fig. 8b von der Annahme ausgegangen, dass die
Signalquellen 119...129 fix installierte Leuchten (entlang
der vom Roboter befahrenen Route) sind, welche die Benutzung der
Loipe auch bei Dämmerung oder Dunkelheit ermöglichen. In
Entsprechung zum primären Zweck der Anwendung eines solchen
Roboters fährt der Trainingsroboter 116 präzise hinter dem
Skilangläufer 117 in dessen Spur her, wobei alle
STQ-Zeitdaten vm-adaptiv erfasst und in dessen TICM- Memory (s. auch
Fig. 5) gespeichert werden. Die Entfernung zwischen Roboter und
Anwender wird durch einen Entfernungs-Sensor exakt geregelt.Um aber
die Antriebs-, Brems- und Lenksysteme des Robotervehikels überhaupt
in Gang bringen zu können, müssten vor der Tour STQ-Zeitdaten in
das TICM geladen werden, die als Referenzdaten dienen könnten.
Daher werden im ersten Schritt die primär erfassten Zeitdaten in den
TICM- Referenz-Datenspeicher übernommen; d.h. aus Tw,Td(1,2,3...)
werden im ersten Schritt Tw',Td'(1,2,3...). In weiteren Schritten
wird das Nachfahren hinter dem Läufer mehrmals mit wechselnder
Geschwindigkeit und Spurverteilung wiederholt - sodass der Roboter
mehr über den Skiläufer "lernt" - worauf immer mehr kovariante
Tw',Td'-Zeitdatenmuster in das Referenz-Datenmemory einströmen, auf
welche die Diskriminator- und Analysatoreinheit des Roboters
zugreifen kann (s. gleichfalls Fig. 5).
Nunmehr setzt das
Interpretations- und Optimierungsprogramm ein, welches nur
"authentische" Tw',Td'- Zeitdaten herausfiltert, die als signifikant
für die beste und effizienteste Linie scheinen, und welches
gleichzeitig die als "irrelevant" erkannten Zeitdaten eliminiert.
Dies kommt einem "Lernprozess" gleich, den das Robotervehikel zu
absolvieren hat, bis es schließlich "autonom skilaufen" kann, d.h.
relativ frei, und nach selbsttätig angeeigneten Mustern und
Selbstentscheidungs-Kriterien; ohne irgendeine Fernsteuerung oder
Regelung durch einen vorprogrammierten Algorithmus. In dieser Phase
fungiert der Trainings-Roboter als "Trainer" oder "Pilot", der die
Aufgabe hat, dem Benutzer zu helfen, ideale
Geschwindigkeitsverläufe, die beste Spur und optimales Timing zu
finden. Die optimale Information, die dem Anwender übermittelt
wird, besteht immer nur aus jener, die vom Roboter selber "erlernt"
worden ist.
Der Trainingsroboter fährt auch bei seiner
"praktischen Arbeit" (d.h. während er dem Anwender behilflich ist)
fort, sich weiter zu verbessern, und seine im TICM gespeicherten
STQ-Referenzdaten fortgesetzt zu optimieren und zu ergänzen. Die
Fähigkeit, Bewegungsverläufe oder externe Signalverläufe und
Objekte zu identifizieren und zu erkennen, ist ausbaufähig. Sie
hängt von der Menge und Vielfalt der benutzten Sensoren und von der
Speicherkapazität des TICM ab. Somit ist es möglich, das
Robotervehikel zu veranlassen, Gefahrenszenarien zu erkennen und
davor akustisch oder optisch zu warnen, sowie Abstände und Zeiten
genauer einzuhalten. Im vorliegenden Anwendungsfall führt das
Vehikel das automatische Spuren und Nachlaufen entlang einer Loipe
sogar dann durch, wenn die Fährte zuvor von einem Schneesturm
verblasen wurde und nicht mehr sichtbar ist. Außerdem hat das
Robotervehikel einen Monitor, auf dem die Eigenbewegung in bezug zur
räumlichen Umgebung visualisiert werden kann, und es verfügt über
elektronische Maßnahmen, um Geschwindigkeiten, Rundenzeiten,
Teilzeiten, Gesamtzeiten oder andere relevante Daten in optischer
oder akustischer Weise auszugeben. Eine wichtige Eigenschaft des
hier gezeigten Roboters besteht auch darin, dass die
Geschwindigkeitsphasen in allen Streckenabschnitten durch einfache
Veränderung (Erhöhung oder Verminderung) des Quellen-Zeittakts
synchron beschleunigt oder verlangsamt werden kann. Diese
Eigenschaft ist beispielsweise erforderlich, um das Tempo des
Trainingsroboters in allen Abschnitten entsprechend der körperlichen
Fitness des Anwenders anzupassen. Dies kann manuell per
Fernbedienung oder automatisch - z.B. mittels Puls- oder
Blutdruck-Datentransponder -
geschehen.
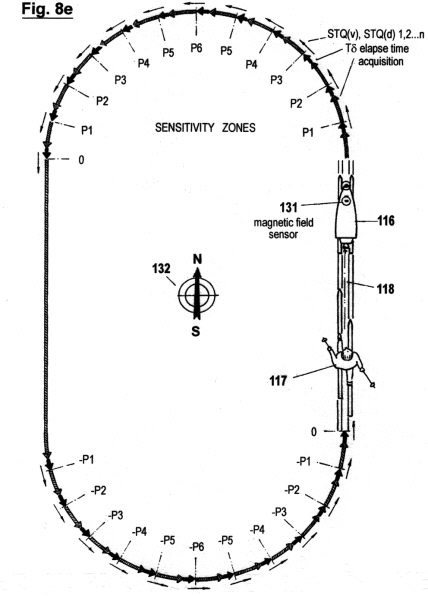
Fig. 8f zeigt die Erfassung von STQ(v)- und STQ(d)-Verstreichzeiten für den Roboter in Fig. 8a, für den Fall, dass Magnetfeld-Sensoren installiert sind. Als Signalquelle wird hierbei das Erdmagnetfeld angenommen. Im gezeigten Beispiel, wo die Spur einen geschlossenen Kreis bzw. eine Schleife bildet, ist die Quantisierung von STQ(i)-Verstreichzeiten uneffizient und daher nicht vorgesehen. Im bezeichneten Bild bewegt sich der Trainingsroboter ("Hase") 116 autonom mit variabler Geschwindigkeit vor dem Ski-Langläufer 117 entlang der Loipe 118. Dabei ergeben sich entlang der Spur unterschiedliche Fahrzeug-Positionen mit variablen Gradienten in bezug zum Erdmagnetfeld 132. Die Magnitude dieser Gradienten wird vom Magnetfeld-Sensor 131 erfasst. Im gezeigten Fall hätte die Magnitude einen sinusförmigen Verlauf. Bei jedem Phasendurchgang zu den Schwellwertzonen P1, P2, P3, P4, P5, P6 usw. werden die besagten STQ(v)- und STQ(d)- Verstreichzeiten adaptiv erfasst und ergeben das aktuelle Td-Zeitdatenmuster, das im TICM gespeichert wird. Die zusätzliche Quantisie- rung von STQ-Verstreichzeiten aus Magnetfeld-Gradienten erleichtert die Ortung kovarianter Tw',Td'-Zeitmuster, die im Referenzdaten-Speicher enthalten sind. Infolgedessen verbessert sich die Autoadaptions- und Erkennungsfähigkeit des Roboter-Vehikels. Je mehr Sensoren in den Autoadaptionsprozess beteiligt sind, desto "autonomer" wird der beschriebene Mechanis- mus (s. auch Fig. 5). In dieser Weise kann ein auf biologisch/chemischen Strukturen basierender selbstorganisierender autonomer Organismus, wie in Fig. 4a - f beschrieben wurde, geschaffen werden.
Fig. 8f zeigt die Erfassung kreisförmiger Positionsfelder mittels GPS-Sensoren. Diese Maßnahme dient (zusätzlich zu jenen in Fig. 8b - e) zur weiteren Verbesserung der zeitlichen und motorischen Auto-Adaption, und zu einem präziseren Autokovarianz-Verhalten und Bewegungs-Nachvollzug. Vorbedingung für eine exakte Funktion ist ein GPS ("global positioning system") von hoher Qualität, die mit äußerst geringen Abweichungen arbeitet. Da in diesem Fall ein rechteckiger Signalverlauf empfangen wird und daher keine Unterteilung in unterschiedliche Empfindlichkeitszonen möglich ist, können nur STQ(v)- und STQ(i)-Verstreichzeiten, nicht jedoch STQ(d)-Verstreichzeiten quantisiert werden (welche - wie gezeigt - zwischen niedrigeren zu höheren Phasendurchgängen und vice versa gemessen werden). In Fig. 8f bewegt sich der Trainingsroboter ("Hase") 116 mit variabler Geschwindigkeit vor dem Skilangläufer 117 entlang der Loipe 118, wobei sich entlang der Spur kreisförmige GPS-Positionsfelder 134a, b...n ergeben, die vom GPS-Sensor 133 mit hoher Messgenauigkeit in reproduzierbarer Weise wahrgenommen werden. Die Radien der Positionsfelder sowie die Trennschärfe zwischen benachbarten Feldern sind justierbar. Bei jeder Detektierung eines neuen Positionsfeldes wird ein Triggersignal an die STQ- Aquisitionseinheit ausgegeben, welches die Registrierung der STQ(v)- und STQ(i)-Verstreichzeiten veranlasst, die dann als aktuelle vm-adaptive Zeitdatensequenzen Tw(1,2,3....) im TICM gespeichert werden. Die Fähigkeit des Roboters zur Optimierung der Auto-Adaption kann gesteigert werden, indem die Anzahl der detektierten Positions-Felder gezählt und verglichen wird, oder indem man den Zeitdaten bei jeder Querung eines Positionsfeldes einen spezifischen Datencode zuordnet.
Fig. 9 ist eine allgemeine schematische Darstellung, und zeigt, wie Zeitdatenströme produziert werden. Jede Amplituden-Querung von Empfindlichkeitszonen oder Schwellwert- potentialen in redundanzarmen autonomen selbstorganisierten Systemen (wie z.B. Organismen oder mechanistische Robotersysteme) führt zur Quantisierung von Verstreich- zeiten, wenn diese über entsprechende Sensoren (oder Rezeptoren) zur Wahrnehmung der externen physikalischen Umgebung verfügen. Diese Basis-Technologie, wie sie im Bild gezeigt wird, behauptet universale Gültigkeit und Anwendbarkeit. Die Darstellung zeigt ein sehr vereinfachtes Schema, das auch vom Nichtexperten klar verstanden wird.
1) Die primäre Funktion jedes Organismus (einschließlich autonomer selbstorganisierender Roboter) besteht in der “Erkundung” seiner Umgebung, um festzustellen, ob zeitlich/räumliche Variation zwischen dem physikalischen Eigenzustand und dem Zustand seiner Umgebung besteht. Dazu dient eine Vielfalt unterschiedlicher Sensoren oder Rezeptoren 135a, b….n usw.
2) Nur wenn eine solche Variation tatsächlich existiert, werden die mit 137a, b…n bezeichneten baktuellen STQ-Verstreichzeiten Tw(1,2,…n ) oder Td(1,2…n)abgeleitet. Die Zeitzählfrequenz zu deren Messung ist abhängig von aktuell erfassten STQ(v)- Quanten Tv(1,2,3…n); 136a, b,c …n welche die Parameter für die zeitlich/räumlichen Abweichungen vm (1,2…n) zwischen Sensoren 135a,b,…n und externen Quellen darstellen. Diese Abweichungen sind mit den “relativen Geschwindigkeiten” vm (1,2..n) ident. Anmerkung: vm (1,2…n ) werden immer mit einer invarianten Zeitzählfrequenz f als Absolut-Zeitbasis (Quellen-Zeittakt) erfasst.
3) Ein Organismus verfügt über “Aktuatoren”, die eine auf sich selbst bezogene aktuelle Veränderung eines zeitlich/räumlichen Zustands (z.B. seine eigene Bewegung) so beeinflussen, das Muster dieser Veränderung weitgehend kovariant ist mit einem früher registrierten Muster einer zeitlich/räumlichen Zustands-veränderung (genauer gesagt: diesem “nacheifert”). Da außerdem bei der Speiche-rung neuer Tw,Td-Sequenzen immer wieder das kürzeste und effizienteste Zeit-muster Priorität hat, versucht der Organismus fortgesetzt, besagte zeitlich/ räumlichen Zustandsveränderungen zu optimieren. Beide Prozesse resultieren ausschließlich aus dem Vergleich von quantisierten STQ-Verstreichzeiten und aus der Erkennung von isomorphen Zeitdatenmustern (s. dazu auch Fig. 5), und werden als “Auto-Emulation” und “Auto-Optimierung” oder aber auch als “Bestreben nach Autokovarianz” bezeichnet.
4) Eine essentielle Konsequenz dieser Betrachtungen besteht darin, dass in allen Organismen der beschriebenen Art eine inhärente teleologische Tendenz zu fortgesetzter Auto-Adaptation und Auto-Optimierung besteht. Dies ruft die Fähigkeit zur Selbstorganisation hervor.
5) Wie aus Fig. 9 eindeutig zu sehen ist, hängen sowohl “Zeit” als auch “Geschwindigkeit” von der Existenz entsprechender Sensorik zu deren Wahrnehmung ab. Genau genommen fließen alle Zeitdaten und Informationen aus der “Gegenwart” (dem Ursprung der Erfassung) in Richtung “Vergangenheit” (der nachweisbaren Existenz). Tatsächlich werden Zeit und Geschwindigkeit nicht als Kontinuum “erfühlt”, sondern in Form von Quanten. Um beide physikalischen Größen als kontinuierlichen Verlauf zu empfinden, erfordert es in einem Organismus enorme Autoadaptions- und Autoemulations-Fähigkeit. Es kann gesagt werden, dass die angeführten fundamentalen Prinzipien nicht nur für Robotik und biologische Einheiten gültig sind, sondern auch für molekulare, atomare und subatomare Strukturen. Auch bei diesen hat es sich um “zeitempfindende Organismen” zu handeln, ansonsten hätten sie keine Basis, um zu existieren. Fazit: Es gibt weder Zeit noch Raum ohne irgendeine andere physikalische Größe ohne subjektive Empfindungsfähigkeit dafür. Objektiv gesehen existiert im Universum ausschließlich Sensorium mit unterschiedlichen Empfindlichkeitsbereichen; und daraus resultiert lokale subjektive Zeitempfindung und ein generelles universelles Bestreben nach Autoadaption, Auto-Optimierung und Auto-Emulation. Dies ist ein fundamentales teleologisches Ordnungsprinzip.
Der Anmelder fasst einige daraus resultierende neue wissenschaftliche Erkenntnisse und Theorien zusammen und postuliert wie folgt:
1) Die hierin beschriebene erfindungsgemäße Methode ist universell anwendbar und zeigt die äußerste (ultimative) Höhe des erreichbaren Stands der Technik auf.
2) Zeitdiskrete Quantisierungsmethoden, in deren Zusammenhang eine empfangene Signalamplitude zu vorbestimmten Zeitpunkten abgetastet und digitalisiert wird, erweisen sich erweisen sich als zu schwach, um daraus hocheffiziente autonome Selbstorganisations-Prozesse zu generieren.
3) In redundanzfreien autonomen selbstorganisierenden Systemen gibt es keine “Zeitpunkte” und keinen Determinismus. In diesen Systemen werden Verstreichzeiten quantisiert, die aus der zeitlich/räumlichen Zustandsveränderung zwischen Sensoren und externen Quellen herleiten.
4) Jedes derartige System verfügt über seinen eigenen Zeittaktgeber und produziert seine eigene Zeit. Die Zeitzählfrequenz zur Quantisierung von Verstreichzeiten wird in auto-adaptiver Weise stets an jene relative Geschwindigkeit vm angepasst, mit der Zustandsveränderungen auftreten.
Die Zeiterfassung hat in jedem Fall Quantennatur, d.h. sie hat die Eigenschaften einer diskreten “Zählung”, egal ob die Erfassung analog, digital oder in sonstiger Weise erfolgt. Außerdem ist die Zeiterfassung subjektiv/passiv, d.h. die Zeitquanten werden “erfühlt” und nicht im konventionellen physikalischen Sinne “objektiv gemessen”.
5) Um imstande zu sein, in autonomen selbstorganisierenden Systemen Verstreichzeiten quantisieren zu können, müssen die einzelnen Sensoren oder Rezeptoren über unterschiedliche Grade (Zonen) von Wahrnehmungsbereichen (oder Schwellwerten) verfügen.
6) Um den Unterschied zwischen “Synchronität” (nach konventionellem Verständnis) und “Auto-Adaption” klarzustellen, sei wie folgt definiert:
a) Parallele Synchronität (kurz: “Synchronität”): d.h. die zeitlichen Änderungen der physikalischen Zustände verschiedener Systeme verlaufen zur gleichen Zeit kovariant.
b) Autonome Adaption (kurz: “Auto-Adaption”): d.h. die zeitlichen Änderungen des physikalischen Zustandes des gleichen Systems verlaufen zu verschiedenen Zeiten kovariant.
7) In allen redundanzfreien autonomen Systemen erhöht sich die Fähigkeit zur Selbstorganisation mit der Menge der dem autonomen Anpassungs- und Optimierungsprozess verfügbar gemachten Verstreichzeit-Parameter sowie mit der Anzahl und der Vielfalt der beteiligten Sensoren bzw. Rezeptoren.
8) Bei “Synchronität” (Definition s. 6a oben) geht im Idealfall die Anzahl quantisierter Verstreichzeitparameter gegen Null, in 6b aber gegen Maximum (wobei außerdem o.g. Pkt. 7 gilt! ). Daher kann davon ausgegangen werden, dass allen autonomen Systemen der genannten Art ein Bestreben nach fortgesetzter Auto-adaption, Auto-optimierung und “motorischer Nach-eiferung” innewohnt. Dieses Bestreben hat eine Affinität zum biologischen Begriff “Lebenskraft”.
9) In autonomen selbstorganisierenden Systemen existiert kein “Timing” (d.h. zeitlich/räumliche Koordination einer Bewegung) ohne den Vergleich aktuell erworbener Verstreichzeitmuster mit früher erworbenen Verstreichzeit-mustern. Kurz gesagt: es gibt kein “Timing” ohne begleitende “Zeitnahme”.
10) Auto-Adaptions-Satz von Erich Bieramperl:
Jede aktuelle nicht-chaotische Zustandsveränderung (A) eines autonomen Systems (X) mit dem variablen dynamischen Verlauf vm(1,2,3..n) unterliegt einer aktuellen Folge von Verstreichzeiten TW(1,2,3 ..n) sowie einer kovarianten Folge von Versteichzeiten TW'(1,2,3..n) aus einer zeitlich versetzten Zustands-veränderung (A’) oder aus einer Kombination unterschiedlich zeitlich ver-
setzter Zustandsveränderungen (A1′)(A2′)..(An’), wonach (A) mit (A’) oder (A) mit (A1)(A2)…(An’) annähernd isomorph ist.
wobei:
TW = vm-adaptiv erfasste aktuelle STQ(i)- oder STQ(d)-Verstreichzeit Tw oder Td und: TW’ = vm-adaptiv erfasste kovariante STQ(i)- oder STQ(d)-Verstreichzeit Tw oder Td
Weitere wissenschaftliche Konsequenzen sind folgende:
11) Jede vorsätzliche Festlegung eines bestimmten Zeitpunktes für eine beabsichtigte Handlung – ein sogenannter “Willensakt” eines autonomen Organismus – resultiert seinerseits aus fortgesetzter autonomer Adaption der beschriebenen Art und ist daher in deterministischer Weise nicht realisierbar.
12) Aus der Fähigkeit eines autonomen Systems, zu aktuell erfassten Verstreichzeitmustern passende früher erfasste Verstreichzeitmuster zu finden – und zu versuchen ihnen nachzueifern – resultieren nicht nur Auto-Adaption, Auto-Optimierung, Selbstorganisation sowie die Erkennung der physikalischen Umgebung und der eigenen Bewegung; sondern letztlich auch Bewegungskoordination, Intelligenz und bewusstes Handeln.
13) Auto-adaptive, auto-optimierende und selbstorganisierende Prozesse der beschriebenen Art haben universellen Anspruch; und zwar nicht nur auf mechanistische Systeme, Roboter, Automaten und biologisch/chemische Organismen, sondern auch auf molekulare und atomare Strukturen. Alle autonomen selbstorganisierenen Systeme enthalten Information in Form von Daten in der Zeitdomäne.
Da in solchen Systemen Zeit “subjektiv empfunden” und nicht “objektiv gemessen” wird (s. Satz 3), folgt weiters:
14) Alle zeitabhängigen physikalischen Größen werden im Universum “subjektiv empfunden”. Gibt es für Zeit und Geschwindigkeit kein entsprechendes Sensorium, so existiert auch in objektiver Weise keine “Zeit”. Beispiel: In “schwarzen Löchern” existiert deshalb keine “Zeit”, weil es kein Sensorium dafür gibt. Das atomare und subatomare Sensorium ist in diesem Fall quasi “tot”.
15) Jede physikalische Zustandsveränderung, der kein auto-adaptiver Prozess zugrunde liegt, verläuft zunehmend chaotisch, woraus folgt, dass die beschriebene Tendenz zu Auto-Adaption im Universum der Tendenz zu Entropie und Chaos (s. zweiter Hauptsatz der Thermodynamik!) entgegen wirkt.
16) Wenn vm zu hoch und STQ(v) zu kurz ist um gemessen (besser: “empfunden”) zu werden, so kann daraus weder ein Auto-Adaptionsprozess (da ja auch keine Verstreichzeiten ableitbar sind) noch ein Selbstorganisationsprozess resultieren. Daher ist z. B. die Lichtgeschwindigkeit c aus jenem Grund eine ultimative Größe, weil sie das kürzeste STQ(v)- Quantum impliziert, das von atomaren Strukturen “wahrgenommen” werden kann.
17) Besteht absolute physikalische Invarianz zwischen dem Sensorium autonomer Systeme und seiner Umgebung, so können ebenfalls keine STQ-Quanten abgeleitet werden. Dies ist die Erklärung dafür, warum beispielsweise der absolute Nullpunkt (-273,15°C) eine ultimative physikalische Größe ist. In diesem Fall ist das atomare und subatomare Sensorium nicht imstande, eine niedrigere Temperatur zu erkennen, weil mangels STQ-Quanten kein Autoadaptions-Prozess mehr stattfinden kann.
18) Wie schon erwähnt, verfügen auch atomare und subatomare Strukturen über sensorische und zeitquantisierende Eigenschaften. Ihre Beschreibung ist aus Sicht der Quantentheorie unzureichend. Gibt es keine Messung oder Beobachtung eines Ereignisses, so existiert weder “Zeit” noch “Geschwindigkeit” (s. auch 13). Quanten-Phänomene, wie sie beim bekannten Doppelspalt-Experiment oder beim SCULLY-Experiment auftreten (“Quanten-Unbestimmtheiten”) sind somit auf diese Weise erklärbar.
19) Elektromagnetische Kräfte, Graviation, starke und schwache Wechselwirkungen (Kernkräfte), sogenannte “Autokatalyse” (KAUFFMAN) , “synergetische Effekte” (H. HAKEN) oder andere Phänomene resultieren aus der Existenz von zeitquantisierendem Sensorium, Autoadaption und Auto-Emulation. Diese Eigenschaften können als inhärentes teleologisches Ordnungsprinzip im Universum betrachtet werden (S. 8).
20) Die Fähigkeit, Zeit und Geschwindigkeit als Kontinuum und nicht als endlose Serie einzelner Verstreichzeiten zu empfinden, erwächst seinerseits aus fortgesetzten Autoadaptions- und Selbstorganisationsprozessen. Je höher die “Intelligenz” eines autonomen Systems als Ergebnis solcher Prozesse, desto ausgeprägter sein subjektives Zeitempfinden und seine Fähigkeit zur Antizipation.
Folgende Konsequenzen ergeben sich daraus für Metamathematik,
Aussagelogik, Erkenntnistheorie und Philosophie:
1) Da es keine “Zeitpunkte” im deterministischen Sinne gibt, kann weder ein Zustand eines Systems zu einem “Zeitpunkt” festgestellt werden, noch können “Zeitpunkte” für künftige Zustände festgelegt werden. Es existiert kein Determinismus irgendeiner Art. Da sowohl die klassische Physik als auch die Quantentheorie auf der Vorbedingung basieren, dass ein System zu einem bestimmten “Zeitpunkt” in einem bestimmten Zustand befindlich ist (im ersten Fall als Punkte des Phasenraums, im zweiten Fall als Wahrscheinlichkeitsverteilungen im Phasenraum) können beide Theorien nicht völlig widerspruchsfrei frei sein. (s. auch
THOMAS BREUER/ 1997)[1]
2) Nach WIGNER (1961)[2] müsste eine absolut universell gültige Theorie imstande sein, auch das Zustandekommen menschlichen Bewusstseins zu beschreiben. Dazu vermag die gezeigte Autoadaptionstheorie imstande zu sein; die Quantentheorie hingegen nicht. (Wigner postulierte, dass komplexe Quantenmechanik nur dort eine brauchbare Beschreibung der physikalischen Realität liefert, wo es kein “subjektives Empfinden” gibt. Der Anmelder vertritt den Standpunkt, dass es subjektives Empfinden auch in atomaren und subatomaren Strukturen gibt.)
3) Verstreichzeitreihen wie TW und TW’ sind als Ketten in einem axiomatischen formalen System betrachtbar; wenngleich es sich dabei um ein “System in der Zeitdomäne handelt und nicht um ein arithmetisches System im Verständnis der klassischen Zahlentheorie. Tatsächlich weist das besagte formale System mindestens ein Axiom auf und leitet durch die Anwendung eines bestimmten Algorithmus fortgesetzt Zahlenketten ab. Nach TURING kann ein axiomatisches zahlen-theoretisches System auch durch eine mechanische Prozedur gegeben sein, welche Formeln und Algorithmen “produziert “.Aus diesem Grund sind daher die bekannten Logik-Theoreme von GOEDEL, TARSKI oder HENKIN auf ein solches Modell durchaus anwendbar. GOEDEL’s Unvollständigkeitssatz [3] zeigt, dass in jedem reichhaltigen zahlentheoretischen Modell widerspruchsfreie Formulierungen enthalten sind, die mit den Regeln desselben Modells nicht bewiesen werden können und demnach unentscheidbar sind. Dies gilt auch für meta-theoretische Modelle und für meta-meta-theoretische Modelle usw. Beispielsweise ist eine selbstbezügliche meta- theoretische Aussage nach Art der Gödel-Formulierung ~ ICH BIN BEWEISBAR – weder beweisbar noch widerlegbar. Ein Entscheidungsverfahren für diese Aussage führt zu einem unendlichen Regress. TARSKI zeigte, dass auch ein Entscheidungsverfahren für zahlentheoretische “Wahrheit” [4] unmöglich ist und in einem unendlichen Regress endet. Somit ist also eine selbstbezügliche Aussage der Art ~ ICH BIN BEWEISBAR – “wahr”, nicht jedoch “beweisbar”. Daraus folgt, dass “Beweisbarkeit” ein schwächerer Begriff ist als “Wahrheit”. HENKIN zeigte, dass es Aussagen gibt, die ihre eigene Beweisbarkeit und “Produzierbarkeit” in einem spezifischen zahlentheoretischen Modell behaupten und demnach unbezweifelbar “wahr” sind [5]. Eine Henkin’s Theorem entsprechende selbstbezügliche Aussage würde etwa so lauten: >es existiert ein zahlentheoretisches Modell, in dem ich beweisbar bin< Ketten von
quantisierten Verstreichzeiten wie TW und TW’ nähern sich dem Geltungsbereich von HENKIN’s Theorem. Würde man Henkin’s Logik darauf anzuwenden, so lautet ihre Aussage etwa: >ich werde entstehen, um bewiesen zu werden<. TW und TW’ sind demnach Ketten oder Aussagen, die in einem spezifischen formalen Modell produziert werden, das sein eigenes Entscheidungsverfahren auf Wahrheit, Wider-spruchsfreiheit, Vollständigkeit und Beweisbarkeit durch fortgesetzte Selbst-Generierung veranlasst (s. dazu auch Beschreibung zu Fig.10).
Im Gegensatz zu selbstbezüglichen Ketten oder Sätzen des Gödel- oder Henkin-Typs behaupten Verstreichzeitketten nie, zu einem gegenwärtigen Zeitpunkt “wahr”, widerspruchsfrei”, “vollständig” oder “beweisbar” zu sein, da jenes “zahlentheoretische Modell”, in dem sie produziert werden, gar keine “Zeitpunkte” kennt. Dieses Modell verbietet auch übergeordnete Semantiken oder Meta-Theorien oder Meta-Meta-Theorien usw. Es ist klar ersichtlich, dass jedes formale System, jede Meta-Theorie, jede Meta-Meta-Theorie und jede Semantik, in der Axiome oder Ketten oder Sätze irgendeiner Art formuliert werden, das Ergebnis fortgesetzter autonomer Adaptation ist (die wiederum auf der Quantisierung von Verstreichzeiten basiert) und somit eine Ableitung aus dem beschriebenen Modell ist.
4) Die Erkenntnis, dass ein spezifisches formales System mit absolutem universellem Anspruch existiert, aus dem alles Seiende hervorgegangen ist und dem alle anderen Systeme unterzuordnen sind, ist nicht neu. Bereits im frühen Altertum (viele Jahre vor PLATO und ARISTOTELES) ließen die Hebräischen Schriften (2. Moses 3-14) den “Quell aller Logik” von sich selbst sagen: “JHWH” (gesprochen: Jahwe oder Jehova), was soviel bedeutet wie: >Ich werde mich als seiend erweisen< [6]. Dieser Satz behauptet also sein eigenes Entscheidungsverfahren auf Beweisbarkeit, Wahrheit, Vollständigkeit und Widerspruchsfreiheit in einem spezifischen formalen System, das er veranlasst, zu “werden”.
5) Es gibt keine “Erkennung” ohne “Wiedererkennung”.
[1] Thomas BREUER (1997) “Quantenmechanik: Ein Fall für Goedel” ISBN 3-8274-0191-7
[2] Eugene WIGNER (1961) “Remarks on the Mind-Body-Question”,
siehe auch: Roger Penrose: Des Kaisers neue Kleider”/ Spektrum-Verlag Heidelberg (S. 287)
[3] Kurt Goedel “Über formal unentscheidbare Sätze der Principia Mathematica und verwandter Systeme I. (1931), siehe auch: Douglas HOFSTADTER “Goedel, Escher, Bach” ISBN 0-394-74502-7 (Seite 19)
[4] Douglas HOFSTADTER “Goedel, Escher, Bach” (s. Seite 618: “Tarski`s Satz”)
[5] Douglas HOFSTADTER “Goedel, Escher, Bach” (s. Seite 577: “Henkin-Sätze”)
[6] Siehe WIKIPEDIA unter JHWH
Fragen dazu bitte richten an: info@sensortime.com